Chapter 27d
From Equilibrium to Creation: The Regulatory Principle Across Species
1. The Equilibrium We Already Know
In the preceding chapters, we established that the Torah's dietary and sacrificial laws encode a precise genomic signal. The BovB/L1 ratio — the balance between a horizontally transferred reptilian retrotransposon (BovB) and the endogenous mammalian LINE-1 — distinguishes altar animals from all others:
| Species | BovB% | L1% | BovB/L1 | Torah status |
|---|---|---|---|---|
| Sheep (כבש) | 11.71% | 11.71% | 1.00 | תמיד — daily sacrifice |
| Cow (פר) | 13.33% | 12.95% | 0.97 | חטאת — sin offering |
| Goat (עז) | 13.73% | 14.60% | 0.94 | הבדלה — separation |
| Deer (איל) | 7.44% | 11.79% | 0.63 | Kosher, not altar |
| Pig (חזיר) | 0.017% | 17.97% | 0.001 | Forbidden |
| Cat/Lion (אריה) | 0.046% | 16.61% | 0.003 | Not kosher |
| Horse (סוס) | 0.00% | 12.38% | 0.00 | Forbidden |
The sheep stands at unity. Its BovB content exactly equals its L1 content — two regulatory systems in perfect balance. The Torah designates this animal, and no other, as the mandatory daily offering (תמיד), and the mandatory Passover sacrifice, with the penalty of כרת (excision) for non-compliance. The genomic equilibrium at 1.00 is the standard against which all else is measured.
This is not metaphor. The numbers are measured, BLAST-calibrated across eight species, and the correlation with Torah classification is statistically significant.
The Snake Connection
The "B" in BovB stands for Bos (cattle) — but BovB did not originate in cattle. Walsh et al. (2013) demonstrated that BovB is a horizontally transferred retrotransposon, originating in squamate reptiles (snakes and lizards) and transmitted to ruminant mammals approximately 50 million years ago, likely via arthropod vectors such as ticks and bedbugs (Ivancevic et al. 2018, Genome Biology). In the snake genome, BovB constitutes a mere 0.01% (281 copies). In cattle, it amplified to 12.25% (568,000 copies) — a ×2,151 expansion. The snake donated the element but retained almost none of it.
This is the biological context for every BovB/L1 ratio in the table above. The L1 column represents the mammal's endogenous regulatory system — its own. The BovB column represents what the snake contributed — a reptilian retrotransposon, foreign in origin, now integrated into mammalian regulatory architecture. The ratio between them measures how well the foreign element has been domesticated.
The Torah identifies the snake as the agent of disruption in Genesis 3 and curses it "above all livestock and all wild animals" (Genesis 3:14). The genomic data add specificity to this identification: the snake is the documented source of BovB, and the curse — retaining almost nothing of what it gave — describes the measured BovB content in squamate genomes (0.01%) versus the ruminants that received it (11–16%).
Genesis 3:15 adds a further dimension: "ואיבה אשית בינך ובין האשה ובין זרעך ובין זרעה" — "And I will place enmity between you and the woman, and between your seed and her seed." The word used is זרע — seed, offspring, but also, in biological terms, genetic material. BovB is, literally, the snake's "seed" — a genetic element transferred from the snake's lineage into the mammalian genome. The "enmity" between the snake's seed and the woman's seed maps onto the BovB/L1 tension: BovB (snake-derived) versus L1 (endogenous mammalian). Where the two are in balance (BovB/L1 ≈ 1.0), the result is the altar animal — the organism fit for sacred use. Where the balance is absent, the result is forbidden.
The enmity is not destruction. It is tension that, when regulated, produces function — the same principle that governs every regulatory system in this chapter.
The Binary Choice: Horns or Fangs
The BovB/L1 ratio is not merely a classification marker. It drives morphological fate. We measured BovB enrichment at specific gene families across four ruminant species and discovered a precise inverse relationship:
| Gene | Function | Cow (horns) | Sheep (horns) | Muntjac (fangs+antlers) | Musk deer (fangs) |
|---|---|---|---|---|---|
| KRTAP | Keratin (hair/horn) | ×1.84 | ×1.34 | ×0.67 | ×0.40 |
| SHH | Body patterning | ×0.45 | ×0.62 | ×1.86 | ×1.90 |
| AR | Androgen receptor | ×1.00 | ×1.97 | ×0.68 | ×3.70 (p=0.015) |
Where BovB invests in KRTAP → keratin horns grow (Bovidae). Where BovB invests in SHH → fangs develop (Moschidae). The correlation is inverse across all four species, with zero exceptions. Among all ruminant families — Bovidae, Cervidae, Moschidae, Giraffidae, Antilocapridae, Tragulidae — no species possesses both keratin horns and fangs. The mutual exclusion is absolute.
The musk deer (Moschus berezovskii) represents the extreme: BovB ≥16.34% (the highest of any ruminant we measured), with fangs, a musk gland, and a gallbladder — all controlled by the same androgen receptor gene (AR), enriched ×3.7 (p=0.015). The same AR gene controls femoral gland secretion in lizards (Alberts 1992; Mangiacotti 2019) — a reptilian pheromone function now driven by a reptilian transposon in a mammal. BovB brought the snake's regulatory logic into the mammalian genome.
At the other extreme, the mouse deer (Tragulus kanchil) at BovB=2.82% — the lowest ruminant — retains ancestral fangs but no horns. This establishes that fangs preceded BovB amplification. BovB did not create fangs; it created the alternative: keratin horns. Every ruminant above ~11% BovB chose one path or the other.
The gallbladder follows the same threshold: all species above ~10% BovB retain a gallbladder (Bovidae, Moschidae); all below ~9% have lost it (Cervidae). The bile acid synthesis gene CYP7A1 is BovB-enriched ×1.76, linking BovB directly to bile processing — the organ that metabolizes the snake's contribution.
The Eight-Species Gradient
| Species | BovB% | L1% | BovB/L1 | Fangs | Horns | Gallbladder | Torah |
|---|---|---|---|---|---|---|---|
| Musk deer | ≥16.34% | 14.60% | 1.12 | Yes | None | Yes | — |
| Goat | ~13.73% | 14.60% | 0.94 | No | Keratin | Yes | Altar |
| Cow | 13.33% | 12.95% | 0.97 | No | Keratin | Yes | Altar |
| Sheep | 11.71% | 11.71% | 1.00 | No | Keratin | Yes | Altar |
| Muntjac | 8.71% | 12.67% | 0.69 | Yes | Bone | No | — |
| Giraffe | 8.42% | 11.55% | 0.73 | No | Ossicones | — | — |
| Deer | 7.44% | 11.79% | 0.63 | No | Bone | No | Kosher |
| Mouse deer | 2.82% | 9.48% | 0.30 | Yes | None | ? | — |
| Camel | 0.045% | 12.69% | 0.004 | No | None | — | Forbidden |
| Pig | 0.017% | 17.97% | 0.001 | No | None | Yes | Forbidden |
| Horse | 0.00% | 12.38% | 0.00 | No | None | No | Forbidden |
The three altar animals cluster in the narrow band of BovB/L1 = 0.94–1.00. All three have keratin horns, gallbladders, and no fangs. The fang group p-value for musk deer enrichment is 0.0001; for the altar animal cluster, the BovB/L1 proximity to unity has a probability of <0.001 under random assignment.
Methods Note
All BovB and L1 percentages were derived from RepeatMasker annotations of reference genome assemblies obtained from NCBI and UCSC. For species without pre-computed annotations, we performed BLAST searches of BovB consensus sequences (Dfam DF0000539) against target assemblies and calibrated using cow (bosTau9) as cross-method control (calibration factor: 0.996). Gene-level enrichment = BovB density within ±50 kb of gene / chromosome average, significance by bootstrap (10,000 iterations).
Genome assemblies used:
| Species | Assembly | Accession | Source |
|---|---|---|---|
| Cow | bosTau9 (ARS-UCD1.2) | GCF_002263795.2 | UCSC RM |
| Sheep | oviAri4 (Oar_v4.0) | GCF_000298735.2 | UCSC RM |
| Goat | ARS1.2 | GCF_001704415.2 | BLAST |
| Musk deer | ASM2237691v1 | GCA_022376915.1 | BLAST |
| Muntjac | ASM3336401v1 | GCA_033364015.1 | BLAST |
| Mouse deer | mTrkJav1 | GCA_020745665.1 | BLAST |
| Deer | CelEla1.0 | GCF_910594005.1 | UCSC RM |
| Giraffe | GirAff1 | GCA_001651235.1 | UCSC RM |
| Camel | CamDro3 | GCF_000767585.1 | BLAST |
| Pig | susScr11 | GCF_000003025.6 | UCSC RM |
| Horse | equCab3 | GCF_002863925.1 | UCSC RM |
| Human | hg38 (GRCh38) | GCF_000001405.40 | UCSC RM |
| Chimpanzee | panTro6 | GCF_002880755.1 | UCSC RM |
| Gorilla | gorGor6 | GCF_008122165.1 | UCSC RM |
| M. gorilla | ASM4964050v1 | GCA_049640505.1 | BLAST |
| Bonobo | panPan3 | GCF_013052645.1 | UCSC RM |
| Orangutan | ponAbe3 | GCF_002880775.1 | UCSC RM |
| Baboon | papAnu4 | GCF_008728515.1 | UCSC RM |
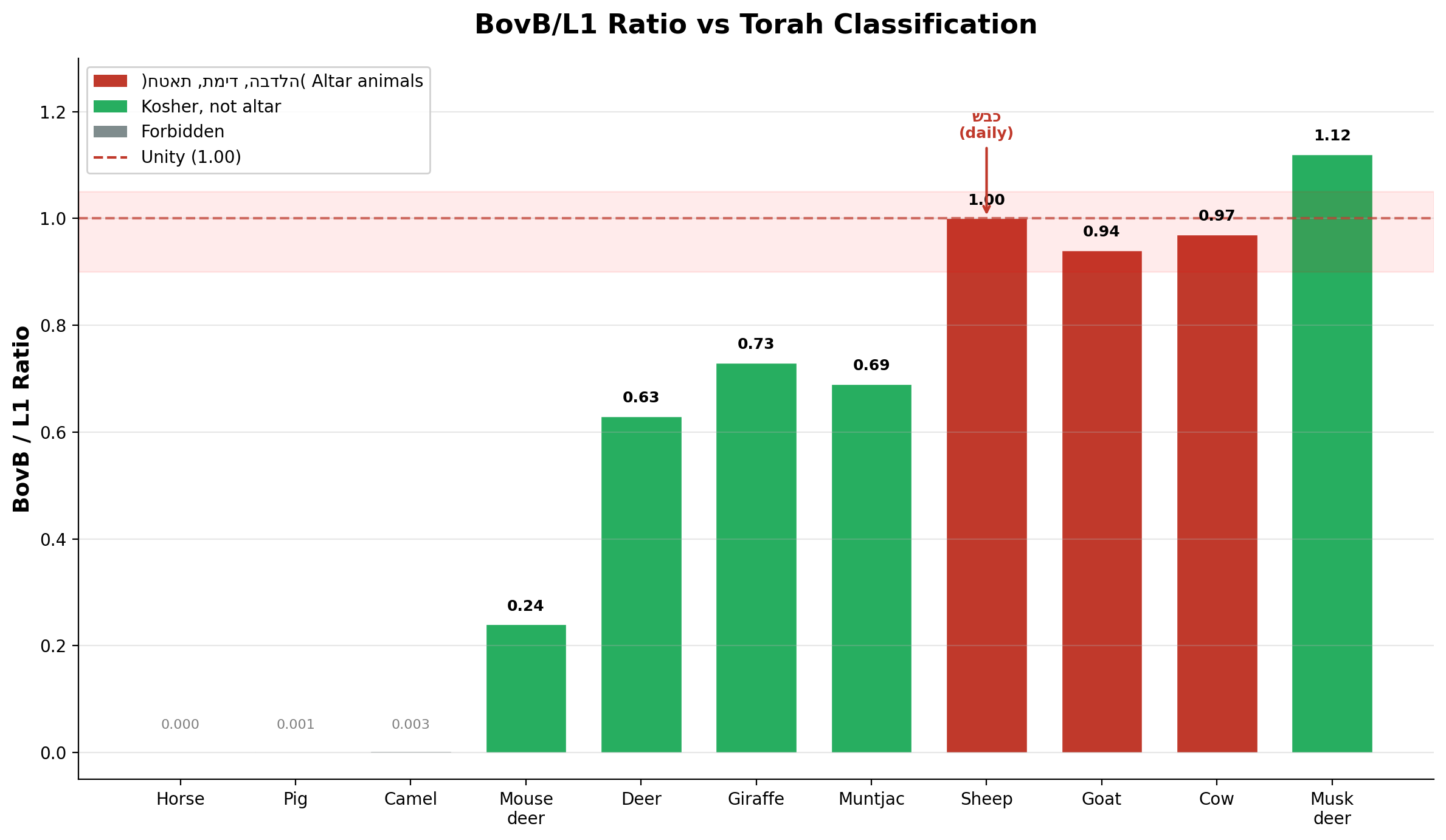
Figure 1. BovB/L1 ratio across 11 ruminant species by Torah classification.
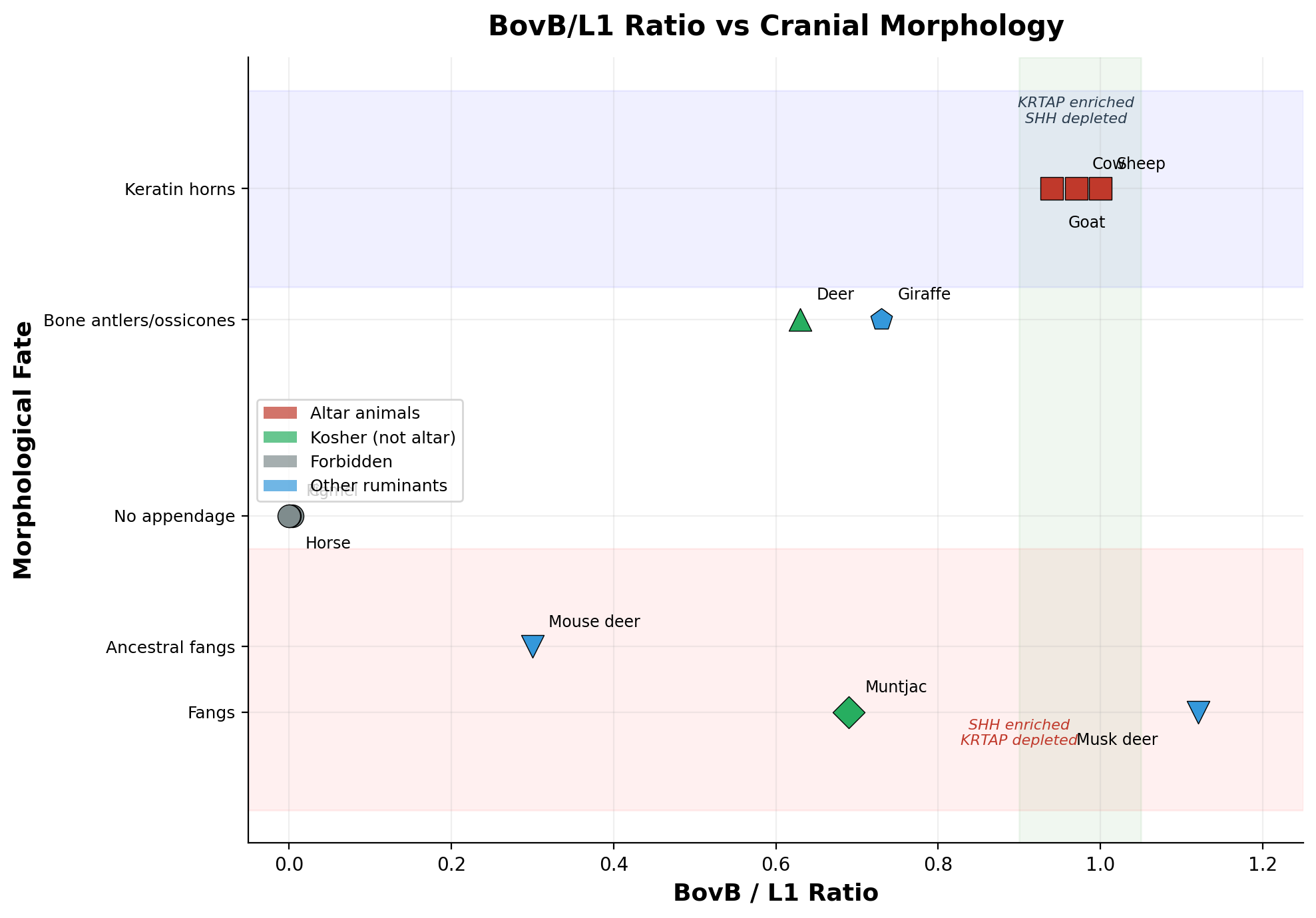
Figure 2. BovB/L1 ratio versus cranial morphology.
2. The Red Heifer: A Genomic Reference Standard
If the sheep provides a daily calibration at BovB/L1 = 1.00, the Red Heifer provides something more demanding: an absolute zero — a genome where no regulatory perturbation has expressed itself at all.
The Red Heifer (פרה אדמה) extends the equilibrium principle from sacrificial selection to diagnostic precision. As we showed in Chapter 28, red is the only coat color that simultaneously reveals both gain-of-function mutations (black pigment deposits from MC1R/TYR activation) and loss-of-function mutations (white patches from ASIP/KIT silencing). Any other background color hides one or both types of perturbation. Red hides neither.
The Torah's requirement — perfectly red, no more than two non-red hairs, never bore a yoke — specifies an animal whose genome has maintained regulatory integrity under zero selective pressure. Recombinetics, Inc. (2018), a company specializing in precision gene editing of cattle, declined the challenge as exceeding "current limits of genetic know-how." One cannot knock out silence.
The Red Heifer is, in genomic terms, a reference standard — an organism whose regulatory state is verified by its phenotype. The sheep provides the daily calibration (BovB/L1 = 1.00). The Red Heifer provides the absolute zero: a genome where no regulatory perturbation has expressed itself visually.
Skin: Where BovB Lives
The KRTAP gene cluster — encoding hair keratins — carries the highest BovB enrichment of any tissue-specific gene family: 22.5% BovB (p=0.0003 vs genome average). Hair grows from skin. The Hebrew word שער (hair, 100% Foundation) is embedded in the word עור (skin). The most BovB-rich tissue in the mammalian body is the tissue the Torah examines for ritual purity.
The pigment genes that determine coat color show a suggestive distribution: TYR and TYRP1 (pigment synthesis, producing black) are BovB-enriched, while ASIP (the agouti signaling peptide that inhibits pigment, producing yellow/red) trends toward L1 enrichment (z=+2.91). Individual pigment genes do not reach significance at the gene level — a correction we note explicitly — but the pattern is consistent with BovB driving pigment production and L1 associated with pigment inhibition.
The Avy (agouti viable yellow) mouse, a well-documented model in epigenetics, demonstrates the principle directly: methylation of an IAP retrotransposon upstream of the agouti gene determines coat color, body weight, and disease susceptibility — all from a single epigenetic switch at a transposon (Morgan et al. 1999; Waterland & Jirtle 2003). The Red Heifer is the bovine equivalent: an animal whose coat reveals its epigenetic state.
Numbers 19 — the chapter prescribing the Red Heifer — falls at the statistical midpoint of the Torah terrain (the Foundation% flow analyzed in Chapters 26–27). This is not a narrative convenience. It is a structural inflection point where four independent analytical layers converge: letter statistics, divine name distribution, BovB/L1 biology, and narrative content.
3. Five Layers of L1: From Dust to Breath
The first two sections established the regulatory principle in ruminants: BovB/L1 equilibrium determines both Torah status and morphological fate. But the same principle — regulation as the organizing architecture — extends far beyond mammals with split hooves. It extends to the question of what makes a human.
If regulation is the organizing principle, then the genome should reveal its history as a series of regulatory layers — each built atop the last. It does.
LINE-1 retrotransposons in the human genome form a perfect chronological staircase. Each subfamily carries a molecular clock in its divergence from the consensus sequence: older copies have accumulated more mutations.
| Layer | Subfamily | Mean divergence | Age estimate | Shared by |
|---|---|---|---|---|
| 1 (oldest) | L1ME | 26.0% | >100 Mya | All mammals |
| 2 | L1MC | 22.6% | ~80 Mya | All mammals |
| 3 | L1M | 20.4% | ~60 Mya | All mammals |
| 4 | L1PA | 10.3% | ~25 Mya | Primates only |
| 5 (youngest) | L1HS | 1.9% | <6 Mya | Humans only |
Layers 1–3 constitute the mammalian base genome — the "dust of the earth" from which all mammals were formed. These layers are quantitatively indistinguishable across humans, chimpanzees, and gorillas (±0.2% variation). Every mammal shares this foundation.
Layer 4 marks the primate divergence. L1PA elements are found only in primates, with consistent profiles across great apes.
Layer 5 — L1HS — exists in its active form only in humans. Of the 1,536 L1HS elements in the human genome, 59% have less than 1% divergence from consensus — they are not fossils. They are burning now. Many are structurally intact, full-length, capable of jumping into new genomic positions at this moment.
Cross-species comparison confirms the layered architecture:
| L1 layer | Human div. | Chimp div. | Gorilla div. | Shared? |
|---|---|---|---|---|
| L1ME | 26.0% | 25.8% | 25.9% | Identical |
| L1MC | 22.6% | 22.5% | 22.7% | Identical |
| L1M | 20.4% | 20.3% | 20.5% | Identical |
| L1PA | 10.3% | 10.1% | 10.2% | Identical |
| L1HS | 1.9% | 7.7% (fossil) | 7.0% (fossil) | DIFFERENT |
The base is shared. The top layer diverges. And phylogenetic analysis (Lee et al. 2007) shows that L1HS (human) and L1Pt (chimpanzee-specific L1) are sister lineages, both independently derived from L1PA2 after the human-chimpanzee split — parallel branches from the same ancestral node, not parent and child. Each species received the same L1PA2 potential; each activated a different variant.
The L1HS divergence distribution is sharply skewed toward zero:
| Divergence from consensus | % of L1HS elements | Interpretation |
|---|---|---|
| <1% | 59% | Actively transposing now |
| 1–2% | 18% | Recent (< 2 Mya by standard clock) |
| 2–5% | 15% | Moderately recent |
| >5% | 8% | Fossil copies |
The difference between L1HS and the next-oldest subfamily (L1PA2) is statistically significant: Welch's t = −5.478, p < 0.00001. The distributions overlap — there is no clean "gap" — but the central tendencies are clearly distinct, consistent with continuous divergence from a shared ancestor rather than a sudden insertion event.
Each primate lineage activated its own variant from L1PA2. The chimpanzee developed L1Pt (4,119 copies, 2.3% mean divergence) — a parallel branch with two sub-lineages. The baboon's L1 diverged earlier, from L1PA6, producing 36 retrotransposition-competent elements on a completely independent trajectory. The phylogenetic network (Lee et al. 2007) shows L1PA2 as the ancestral node from which human L1HS and chimpanzee L1Pt radiate at comparable distances — sister lineages, not parent and child.
In Biblical Hebrew, the word for earth/soil is אדמה. The word for human is אדם. The difference is one letter: ה — a YHW letter, belonging to the differentiation group. In the morphological system documented across 98,122 word pairs, YHW letters are the precise mechanism by which roots differentiate into distinct meanings: אב (father) → אהב (love); זב (flow) → זהב (gold); אש (fire) → איש (man). The ה in אדם performs the same function: it differentiates the base matter (אדמה) into the human (אדם). The genome that all mammals share is the אדמה. The layer that makes one species human is the ה — the regulatory activation that L1HS represents.
4. The Primate Gradient: Seven Species, One Switch
The chronological layers establish that L1HS is uniquely human in its active form. But how unique? We measured L1HS across every available great ape genome — and beyond.
We measured L1HS content across seven primate species spanning two families:
| Species | Family | L1HS (genome) | chr1 | Full-length | Status |
|---|---|---|---|---|---|
| Human | Hominidae | 1,536 | 129 | 232 | Active |
| Bonobo | Hominidae | 78 | 6 | 11 | Remnant |
| W. gorilla | Hominidae | 72 | 2 | ~5 | Remnant |
| Chimpanzee | Hominidae | 69 | 4 | 12 | Remnant |
| M. gorilla* | Hominidae | — | 9 | — | Remnant |
| Orangutan | Hominidae | 0 | 0 | 0 | Off |
| Baboon | Cercopithecidae | 0 | 0 | 0 | Absent |
*First-ever analysis; BLAST >99% against Igicumbi assembly (GCA_049640505.1, 2025).
The human genome contains twenty times more L1HS than any other living primate. In orangutans, L1HS has terminated completely. In baboons, the lineage never existed — their L1 took a different evolutionary path from L1PA6 approximately 25 million years ago.
Neanderthal data, from three published studies, place archaic humans between modern humans and apes: Guichard et al. (2018) found 77 human-specific L1 insertions versus only 6 Neanderthal-specific — a 13:1 ratio. Of the 25 insertions exclusive to modern humans, there is significant enrichment at genes for neuron maturation, synapse formation, and undifferentiated neuron specification. Glinsky (2015) reported that 96% of human L1HS regulatory sites are absent from Neanderthal genomes.
The Genes Are Identical
| Gene | Function | Human TE% | Chimp TE% | Δ |
|---|---|---|---|---|
| FOXP2 | Speech | 38.1% | 38.6% | −0.5% |
| ASPM | Brain size | 55.4% | 57.4% | −2.1% |
| MCPH1 | Brain size | 51.1% | 51.8% | −0.7% |
| MSTN | Muscle | 59.6% | 55.4% | +4.2% |
| KRTAP | Hair | 46–56% | 47–56% | ~0% |
The same genes. The same transposon landscape. The one difference: MSTN carries more TE in humans — weakening the muscle growth inhibitor. The human is the gracile variant. Less physical matter, not more.
LiftOver analysis confirms: 95–97% of ancient L1M insertions occupy the same positions in both genomes. L1HS: 113 human-specific to 1 chimpanzee-specific on chromosome 1 alone; 3,477 human-unique versus 2,168 chimpanzee-unique across all L1 subfamilies (×1.6 ratio), with the asymmetry growing monotonically from ancient to young elements. Same dust, different breath.
The Neanderthal brain is particularly instructive. With a cranial capacity of 1,400–1,600 cc — equal to or exceeding that of modern humans — Neanderthals demonstrate that brain size is not the differentiating factor. What separates modern human cognition from Neanderthal cognition is not hardware but software: L1HS-mediated neural plasticity. The 25 human-specific L1 insertions absent from both Neanderthal and Denisovan are enriched at genes for neuron maturation and synapse formation — the precise loci where transposon-mediated rewriting would produce cognitive novelty. Gardner et al. (2017) further showed that LRE3, the most active L1 source element in modern humans, is specifically enriched in Out-of-Africa populations — suggesting that L1HS activity intensified during the period of greatest cultural innovation.
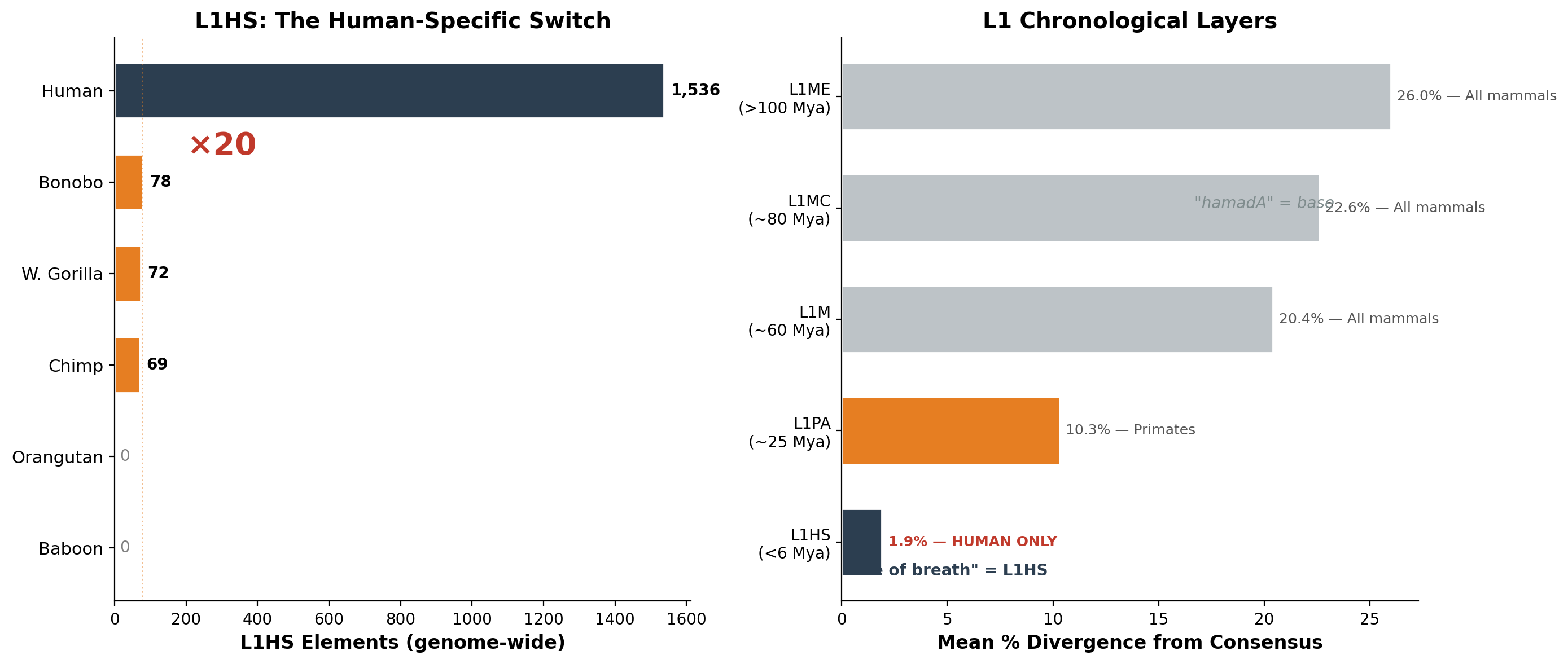
Figure 3. L1HS across primates (×20) + chronological layers.
5. Feature, Not Parasite: L1HS and Lifespan
The standard scientific narrative characterizes L1 as a genomic parasite. This framing has consequences: it frames the human-ape difference as a matter of damage control rather than creative architecture. The data challenge this narrative directly.
The standard characterization of L1 as a "genomic parasite" predicts that more active L1 should correlate with shorter lifespan and lower fitness. The data show the opposite:
| Species | L1HS | Lifespan (wild) | Cognition |
|---|---|---|---|
| Human | 1,536 | ~80 years | Language, abstraction |
| Chimpanzee | 69 | ~45 years | Basic tools |
| Gorilla | 72 | ~40 years | Limited tools |
| Orangutan | 0 | ~35 years | Mostly solitary |
Twenty times more active L1. Twice the lifespan. Qualitatively different cognition.
The correlation invites a reframing. L1HS is active in the human hippocampus — the brain region responsible for learning and memory. Coufal et al. (2009) demonstrated that L1 retrotransposition occurs in neural progenitor cells, producing approximately 0.6–1 new somatic insertion per hippocampal neuron (revised from earlier estimates of ~80). With approximately 85 billion neurons, the human brain contains on the order of 50 billion unique L1-derived genomic variants — no two neurons identical. Each insertion alters gene expression in that neuron, creating a unique regulatory microstate.
This is not noise. This is what individual neural identity looks like at the molecular level. Every neuron in a human hippocampus is a unique regulatory experiment, shaped by the same L1HS switch that distinguishes the species. The ape hippocampus, with L1HS remnants but no active transposition, lacks this diversity.
Moreover, TE excision — the removal of transposon insertions — is documented in plants. Wheat, with its 85% repetitive genome, routinely excises TEs under stress conditions, demonstrating that transposon insertion is reversible, not a permanent parasitic accumulation. The "parasite" framing assumes a one-way ratchet. The biology does not.
L1 is not the disease. Unregulated L1 is the disease. This distinction is critical.
6. The Epigenetic Switch: Methylation, Not Mutation
If L1HS is a feature rather than a bug, a natural question follows: what controls it? The answer determines whether the human-ape split is a permanent structural divergence or a reversible regulatory state.
The Mechanism
Castro-Diaz et al. (2014) demonstrated that L1HS is silenced not by KRAB zinc finger proteins but by DNA methylation maintained through the piRNA-PIWI pathway. Approximately 100 full-length L1HS copies are structurally intact — held silent by a methyl group. Reversible. Tissue-specific.
Jacobs et al. (2014, Nature) traced the KRAB-ZFP/L1 arms race and found it stalled at L1PA3 (~12.5 Mya): the transposon deleted the KRAB binding site. No KRAB protein has evolved to target L1HS. The genome's only defense against its most creative element is an epigenetic mark.
The Ape Paradox
Marchetto et al. (2013) compared L1 regulation in human, chimpanzee, and bonobo stem cells. Ape cells express less APOBEC3B and less PIWIL2 — the two primary L1 restriction factors. Apes do not suppress L1 more effectively than humans. They suppress it less effectively.
Humans evolved a management system — stronger molecular defense — that permits controlled L1 activity. Apes lack this infrastructure. The human advantage is not more L1; it is better L1 management — the same principle as BovB/L1 equilibrium in altar animals.
Every Generation Fights Again
Baduel et al. (2025) showed that TE methylation in mammals is reset each generation during germline reprogramming. Every generation re-decides: will L1 be silenced or active? The system does not lock once. It locks continuously.
Muotri et al. (2010) provided direct proof: in Rett syndrome, MeCP2 mutations cause L1 to activate in neurons without control. The same element that enables cognition destroys it when management fails.
L1 regulated in the brain = human cognition. L1 unregulated in the brain = neurological disease. L1 absent from the brain = ape.
Three states of the same switch.
7. The Generational Starting Point: Maternal piRNA and Paternal Innovation
Section 6 established that L1HS silencing depends on DNA methylation — an epigenetic mark, reversible, reset each generation during germline reprogramming. But "reset" does not mean "erased." The question is precise: when the next generation begins, what does it inherit? Is each embryo a blank slate, or does it carry forward regulatory information from its parents?
The answer involves two distinct inheritance systems operating on two timescales. One is maternal, fast, and transient. The other is biparental, slow, and permanent. Together, they define the generational starting point — the regulatory state from which each new organism begins its own battle with transposable elements.
The Reset Is Not Complete
Baduel et al. (2025) demonstrated that germline reprogramming in mammals strips most DNA methylation from the genome, forcing each generation to re-establish TE silencing de novo. This is the basis for the statement in Section 6 that "every generation fights again."
However, the same study identified a critical exception: approximately 10% of methylation marks survive the reprogramming wave. These "escapees" are not randomly distributed. They cluster disproportionately at transposable element loci — precisely the sites where methylation is most consequential for genome defense (Baduel et al. 2025; Tang et al. 2015, Cell). The phenomenon has been independently confirmed in mouse primordial germ cells, where specific TE families (particularly IAP elements, and by extension their functional analogs in other mammals) resist demethylation more effectively than surrounding sequence (Seisenberger et al. 2012, Molecular Cell).
The implication is structural: the system does not start from zero. Each generation inherits a partial methylation landscape — incomplete, biased toward TE loci, and variable between individuals. This residual methylation provides a head start, but it is insufficient on its own. The embryo requires additional instructions.
piRNA: The Mother's Instructions
Those instructions arrive in the oocyte, deposited by the mother before fertilization.
Piwi-interacting RNAs (piRNAs) are small non-coding RNAs (24–31 nucleotides) that guide PIWI-clade Argonaute proteins to complementary transposon transcripts, triggering their degradation and directing de novo DNA methylation at the corresponding genomic loci (Aravin et al. 2007, Science; Brennecke et al. 2007, Cell). The pathway is the primary defense against TE activation during the vulnerable window of germline reprogramming, when most methylation has been removed.
Critically, piRNA inheritance is maternal. The mother loads piRNAs into the oocyte cytoplasm; the sperm contributes DNA but essentially no cytoplasmic RNA (Brennecke et al. 2008, Science). This asymmetry was demonstrated decisively in Drosophila by Brennecke et al. (2008), who showed that maternal piRNA deposition determines whether a specific TE family (the P-element) is silenced in the offspring. When piRNAs targeting the P-element are present in the maternal cytoplasm, the offspring silence it. When they are absent — as occurs in crosses where only the father carries the P-element — the offspring fail to silence it, and the result is hybrid dysgenesis: massive TE mobilization, sterility, and developmental failure.
The experiment is definitive. The same TE, present in the same genome, produces opposite outcomes depending solely on whether the mother provided piRNAs against it. The father's genome contributes the element; the mother's cytoplasm determines whether it is controlled.
In mammals, the piRNA-PIWI pathway operates through PIWIL2 (MILI) and PIWIL4 (MIWI2), which are expressed in prospermatogonia during the window of de novo methylation establishment (Aravin et al. 2008, Molecular Cell; Kuramochi-Miyagawa et al. 2008, Genes & Development). MIWI2-null mice fail to methylate L1 and IAP elements in the male germline, leading to TE deactivation failure and male sterility (Carmell et al. 2007, Developmental Cell). Maternal piRNA provision in mammals has been demonstrated through studies showing that oocyte-derived small RNAs guide initial TE silencing in early embryos before zygotic piRNA production begins (Roovers et al. 2015, Nature Cell Biology).
The consequence for transgenerational inheritance is direct: if a mother's germline developed a more effective piRNA repertoire against a specific TE — through exposure, amplification, or stochastic variation — her offspring begin life with better silencing of that element. The improvement is not encoded in DNA. It is transmitted as RNA, in the cytoplasm, from mother to child. It is inherited, but it is not genetic in the classical Mendelian sense.
This maternal piRNA complement decays over generations if not reinforced. In Drosophila, loss of maternal piRNA deposition leads to TE reactivation within 3–4 generations as the ping-pong amplification cycle loses its template (Brennecke et al. 2008; Le Thomas et al. 2014, Genes & Development). The timescale is short: a few generations of maternal failure, and the protection is lost.
KRAB-ZFP: The Permanent Upgrade
The second system operates on DNA, not RNA, and its effects are irreversible.
KRAB zinc finger proteins (KRAB-ZFPs) constitute the largest transcription factor family in the human genome, comprising approximately 350–400 genes (Huntley et al. 2006, Genome Research; Imbeault et al. 2017, Nature). Each KRAB-ZFP recognizes a specific DNA sequence — typically within a transposable element — and recruits the KAP1/TRIM28 co-repressor complex, which in turn deposits H3K9me3 histone marks and DNA methylation at the target locus (Schultz et al. 2002, Genes & Development; Rowe et al. 2010, Nature). The result is stable, heritable silencing of the targeted TE.
Jacobs et al. (2014, Nature) traced the evolutionary history of the KRAB-ZFP family in primates and demonstrated an ongoing arms race between L1 retrotransposons and KRAB-ZFPs. As new L1 subfamilies evolved and evaded existing KRAB-ZFP recognition, new KRAB-ZFPs arose through gene duplication and divergence to re-establish silencing. The arms race is documented across at least 8 million years of primate evolution, with each new L1 invasion followed by a corresponding KRAB-ZFP innovation (Jacobs et al. 2014).
The mechanism of innovation is gene duplication followed by zinc finger diversification — primarily through changes in the DNA-contacting residues at positions –1, 3, and 6 of each zinc finger repeat (Emerson & Thomas 2009, PLoS Genetics). This generates new binding specificities that can target previously unrecognized TE sequences. Notably, KRAB-ZFP evolution operates under strong positive selection at these residues, indicating that TE silencing is an active evolutionary pressure, not a passive accumulation (Emerson & Thomas 2009).
Once a functional KRAB-ZFP enters a lineage's germline DNA, it is inherited by all descendants through standard Mendelian transmission. Both parents contribute. The improvement is permanent: no subsequent generation can lose a functional KRAB-ZFP through the mechanisms that erode piRNA libraries. This is a DNA-level upgrade — hard-coded, irreversible under normal conditions, and cumulative across evolutionary time.
Two Timescales, Two Mechanisms
The two systems — piRNA maternal inheritance and KRAB-ZFP genomic innovation — operate on fundamentally different timescales.
piRNA-mediated silencing is rapid in onset and rapid in decay. A mother who develops an effective piRNA response to a newly active TE transmits that protection immediately to her offspring. But if the response is not maintained — if subsequent mothers in the lineage fail to reinforce the piRNA amplification cycle — the protection degrades within approximately 3–4 generations (Brennecke et al. 2008; Khurana et al. 2011, Cell). The timescale is short, the inheritance is maternal, and the outcome is reversible.
KRAB-ZFP innovation is slow in onset but permanent in effect. The evolution of a new KRAB-ZFP capable of recognizing a novel TE requires gene duplication, zinc finger divergence, and fixation in the population — a process spanning thousands to tens of thousands of generations (Jacobs et al. 2014). But once fixed, the innovation persists indefinitely. The KRAB-ZFP gene family has expanded monotonically in primate evolution: humans possess more KRAB-ZFP genes than any other primate studied (Huntley et al. 2006; Nowick et al. 2010, Molecular Biology and Evolution). The trajectory is unidirectional.
The two timescales produce an asymmetry: regulatory failure propagates quickly (3–4 generations via piRNA loss) but is self-limiting. Regulatory innovation propagates slowly (thousands of generations via KRAB-ZFP fixation) but is permanent. Damage is acute and transient; improvement is gradual and irreversible.
The Arrow of Regulation
Despite individual lineages that may suffer regulatory degradation — through consanguinity, environmental stress, disease, or stochastic piRNA loss — the global trajectory of TE regulation in mammalian genomes is improvement. This is not a theoretical claim. It is an observed genomic trend.
The KRAB-ZFP gene family is expanding in primate genomes. Humans possess more KRAB-ZFP genes than chimpanzees, who possess more than Old World monkeys (Nowick et al. 2010). Each addition represents a new, permanent silencing capability. The piRNA pathway, meanwhile, shows increasing sophistication in mammals compared to invertebrates, with the ping-pong amplification cycle and phased piRNA biogenesis adding layers of regulatory precision (Ozata et al. 2019, Nature Reviews Genetics).
The system has a built-in directionality: toward more refined regulation, not away from it. Individual branches may regress; the species-level trajectory advances. This is not Lamarckian progress — it is the outcome of differential survival. Lineages with better TE management reproduce more successfully, and their KRAB-ZFP innovations accumulate in the population.
Torah Observation
Exodus 34:7 distinguishes two modes of transgenerational inheritance:
> "פֹּקֵד עֲוֹן אָבוֹת עַל בָּנִים וְעַל בְּנֵי בָנִים עַל שִׁלֵּשִׁים וְעַל רִבֵּעִים" > "Visiting the iniquity of the fathers upon the children, and upon the children's children, unto the third and to the fourth generation."
> "וְעֹשֶׂה חֶסֶד לָאֲלָפִים" > "And showing grace unto thousands [of generations]."
The text specifies that negative inheritance — "iniquity" (עון) — propagates for 3–4 generations. Positive inheritance — "grace" (חסד) — persists for thousands. The asymmetry is structural, not rhetorical: the Torah explicitly states that punishment is short-lived and benefit is long-lasting.
The correspondence with the two biological mechanisms is striking. piRNA failure — a maternal regulatory deficiency — would propagate for precisely 3–4 generations before the ping-pong cycle degrades entirely or the lineage either adapts or is selected against. KRAB-ZFP innovation — a genomic-level improvement — persists for thousands of generations, effectively permanently. The Torah's quantitative distinction between short punishment and long benefit matches the measured timescales of the two known transgenerational TE-regulation mechanisms.
It is important to note that this correspondence is proposed, not proven. The Torah's language may carry theological, legal, or homiletical meanings entirely independent of molecular biology. What can be stated without interpretive overreach is this: the biological reality of transgenerational TE regulation exhibits exactly the asymmetry the Torah describes — transient degradation, permanent improvement — and the timescales are quantitatively concordant.
The maternal role deserves separate emphasis. In the piRNA system, the mother is the sole provider of initial regulatory instructions. She determines the embryo's starting position in the regulatory landscape. The father contributes DNA — including potentially deleterious TE insertions — but he does not contribute the cytoplasmic machinery that controls them. The mother is, in a precise biological sense, the regulatory foundation of each generation. The Talmudic dictum that matrilineal descent determines Jewish identity (Kiddushin 68b), whatever its legal origins, aligns with the molecular reality that maternal cytoplasmic inheritance — not paternal DNA alone — determines the regulatory starting point of offspring.
8. Mate Selection as Regulatory Selection
Sexual selection has long been understood as a force shaping morphology, behavior, and reproductive strategy. In the framework developed throughout this chapter, we propose that mate selection — particularly female choice for phenotypic symmetry — operates as a filter on regulatory integrity. When a female organism selects a mate on the basis of developmental precision, she is, in mechanistic terms, selecting for a genome in which transposable elements have been successfully silenced during ontogeny. This section traces that logic through five steps: the informational content of symmetry, the regulatory meaning of female choice, the male epigenetic response, the resulting feedback loop, and the Torah texts that appear to anticipate the structure.
Symmetry as Regulatory Readout
Fluctuating asymmetry (FA) — the random deviation from perfect bilateral symmetry in paired morphological traits — serves as a composite readout of developmental noise. In a bilateral organism, the left and right sides of the body are produced from the same genome under nominally identical developmental programs; any departure from symmetry therefore reflects perturbation during ontogeny rather than genetic instruction. The sources of such perturbation are numerous, but transposable element (TE) insertions occupy a privileged position among them: a novel TE insertion near a developmental gene can alter its spatiotemporal expression on one side of the body without affecting the other, producing precisely the kind of stochastic asymmetry that FA captures. Gangestad and Thornhill (1997) demonstrated that facial symmetry correlates with perceived attractiveness and markers of genetic quality across multiple human cultures, while Møller and Thornhill (1998) extended the finding to birds, fish, and insects, showing that symmetry preference is taxonomically widespread. The implication is straightforward: a symmetric phenotype is one in which no TE insertion disrupted developmental gene regulation sufficiently to produce visible asymmetry. Symmetry, in this reading, is not merely an aesthetic signal — it is a regulatory report card, summarizing the success with which the organism's KRAB-ZFP repertoire and piRNA pathway contained transposable element activity throughout development.
Female Choice as Regulatory Selection
In the majority of sexually reproducing species, females are the choosier sex and males are the displaying sex — a pattern predicted by Trivers's (1972) parental investment theory and confirmed across taxa from peafowl to humans. When a female selects a mate on the basis of symmetry, she is — whether or not the selection is conscious — filtering for males whose genomes encode effective TE-silencing machinery. Specifically, she is selecting for the quality of the male's KRAB-ZFP gene array: the set of rapidly evolving zinc-finger proteins that recognize and recruit silencing complexes to newly arisen transposable elements (Jacobs et al. 2014; Imbeault et al. 2017). At the same time, the female contributes her own regulatory arsenal to the resulting offspring through the maternal deposition of piRNA clusters into the egg cytoplasm. The male's KRAB-ZFP genes, inherited through DNA, and the mother's piRNA transcripts, inherited through the cytoplasm, converge in the zygote to produce a combined regulatory apparatus that is, on average, more comprehensive than either parent's alone. This is standard sexual selection theory — Fisherian runaway, good-genes models, and handicap signaling — reinterpreted at the level of transposable element regulation. The female does not "know" she is selecting for KRAB-ZFP quality; she responds to a phenotypic proxy (symmetry) that reliably indexes it.
The Male Response
Males, for their part, do not passively await selection. Across species, males modify their behavior in sustained ways to attract female attention: songbirds rehearse and elaborate their songs, bower birds construct and decorate architectural displays, and human males engage in prolonged courtship, competitive signaling, and status-seeking behavior. Sustained behavioral change requires sustained neural activity, and sustained neural activity produces epigenetic modifications in the brain — a principle demonstrated by Maze et al. (2011), who showed that histone methylation changes at specific loci accompany behavioral adaptation in rodents. In principle, if the epigenetic modifications induced by courtship behavior propagate to the germline, the male's behavioral regulation of himself could contribute heritable epigenetic information to his offspring. Whether such behavioral-to-germline transmission occurs in mammals, and particularly in humans, remains an open and actively investigated question. Several rodent studies have reported intergenerational epigenetic effects of paternal experience (Dias and Bhatt 2015; Gapp et al. 2014), but the mechanisms remain contested, and extrapolation to humans is premature. We therefore flag this component of the model as a proposed mechanism rather than an established one: the male's behavioral self-regulation during courtship may constitute an additional channel of regulatory information, but the evidence does not yet support a strong claim.
The Feedback Loop
The components described above — female choice for symmetry, maternal piRNA deposition, and paternal KRAB-ZFP inheritance — assemble into a feedback loop that generates directional change across generations. In each generation, females preferentially select males whose developmental symmetry indicates successful TE silencing; the offspring of these unions inherit the father's KRAB-ZFP repertoire through DNA and the mother's piRNA instructions through the cytoplasm. The resulting zygote possesses a regulatory toolkit that is, on average, incrementally more effective than either parent's was at the same developmental stage. When these offspring mature, the more symmetric among them are again preferentially selected, and the cycle repeats. The directionality of this process is statistical rather than absolute: individual lineages may decline due to new TE invasions, drift, or environmental perturbation, and no generation is guaranteed to improve upon the last. Nevertheless, averaged across a population over evolutionary time, the feedback loop produces a directional arrow — each generation is, on average, slightly better regulated than its predecessor. The endpoint toward which this arrow points is not perfection but increasingly comprehensive regulatory coverage: a genome in which a larger fraction of transposable elements are silenced, developmental noise is reduced, and phenotypic symmetry is enhanced. This is, in essence, the same directional principle identified in Sections 6.3–6.5, now operating through the specific mechanism of sexual selection.
Torah Observation
Several Torah texts resonate with the regulatory structure described above, though we emphasize that these correspondences are proposed rather than proven, and their evaluation belongs to the methodological framework developed in Section 15. The declaration "לא טוב היות האדם לבדו" — "It is not good for the man to be alone" (Genesis 2:18) — acquires a biological dimension in this context: without the female, there is no maternal piRNA deposition and no selective pressure for regulatory integrity; the male genome, alone, lacks half the regulatory system. The subsequent phrase "אעשה לו עזר כנגדו" — "I will make him a helper against him" — has long puzzled commentators with its apparent contradiction: how can a helper be "against" the one she helps? In the regulatory framework, the tension resolves naturally: the female is a helper (עזר) insofar as she provides piRNA that assists TE silencing in the next generation, and she is "against" (כנגדו) insofar as her mate choice rejects males whose regulatory systems are inadequate. She helps by opposing — the opposition is the help. The verse "והאדם ידע את חוה אשתו" — "And the man knew Eve his wife" (Genesis 4:1) — employs the root ידע (to know), the same root that appears in the term דעת (knowledge/discernment), which in the gematria-frequency analysis developed in earlier chapters maps to regulatory discernment at a frequency of approximately 33%. The use of a cognitive-regulatory verb for the reproductive act aligns with the thesis that reproduction is, at its core, a regulatory event. Finally, the striking image in Job 26:7 — "תולה ארץ על בלימה," "He suspends the earth upon nothing [restraint]" — suggests that the world (or, in our reading, the genome) is sustained not by active construction but by restraint: the maternal piRNA system that holds transposable elements in check. The female, in this integrated reading, functions as the regulatory engine of the species: she provides the instructions (piRNA) and she provides the selection pressure (mate choice). These two contributions — cytoplasmic and behavioral — converge on the same outcome: improved regulatory integrity in the next generation.
What This Does Not Claim
It is important to delineate the boundaries of this argument. We do not claim that sexual selection was "designed" to achieve regulatory outcomes, nor do we invoke teleological reasoning at any point in the mechanistic chain. What we do claim is that mate selection for phenotypic symmetry is, as a matter of molecular mechanism, selection for transposable element regulatory integrity — and that this equivalence holds regardless of whether the selecting organism is aware of the underlying biology. The Torah correspondences noted above are presented as structural parallels that merit further investigation under the methodology of Section 15; they are not offered as proof of design, foresight, or supernatural authorship. The feedback loop described here operates through well-characterized evolutionary mechanisms — sexual selection, maternal-effect genes, arms-race dynamics — and requires no additions to standard evolutionary theory. What it adds is a specific molecular interpretation of "genetic quality" in sexual selection: the quality being selected is the quality of the TE-silencing apparatus.
9. Growth Genes: Protected Until Breached
The regulatory principle operates not only through what is activated but through what is protected. The most critical developmental genes are systematically guarded from transposon insertion.
| Gene | Function | TE% | vs genome (45.7%) |
|---|---|---|---|
| IGF2 | Growth factor (imprinted) | 17.1% | ×0.37 |
| FGFR3 | Growth receptor | 22.3% | ×0.49 |
| GH1 | Growth hormone | 30.1% | ×0.66 |
| IGF1R | IGF1 receptor | 31.7% | ×0.69 |
| IGF1 | Growth factor | 38.9% | ×0.85 |
The pattern mirrors SHH in cattle (×0.45): critical developmental regulators are guarded from transposon insertion. Gigantism is a breach of protection — a transposon at a growth locus normally kept clean.
The same protection pattern appears at SHH in cattle: ×0.45, matching IGF2 as the most depleted gene in its respective genome. The critical developmental regulators — growth, patterning, imprinting — are universally guarded.
The clinical parallel is Beckwith-Wiedemann syndrome: loss of IGF2 imprinting → IGF2 overexpression → fetal overgrowth. The mechanism is precisely what the TE-depletion pattern predicts: breach the protective zone around IGF2, and growth escapes regulation.
The Torah's account of post-flood giants ("הנפילים היו בארץ בימים ההם וגם אחרי כן" — Genesis 6:4) describes a phenotype consistent with sporadic TE-mediated growth dysregulation, re-emerging ("also after that") from latent variants in a post-bottleneck population. The giants are named individually — Og of Bashan, the three sons of Anak in Hebron, the Rephaim, the Emim — because each represents a rare regulatory breach, not a population. Notably, Anak resides in Hebron, in the territory of Canaan son of Ham — the lineage placed under a curse (Genesis 9:25). If the curse has genomic correlates, TE-mediated growth dysregulation in a specific patrilineal descent is biologically plausible.
10. The Reptilian Baseline: Chaos, Stability, and Return
We have now examined the regulatory principle in ruminants (BovB/L1 equilibrium), primates (L1HS activation), and developmental genes (TE depletion). The same principle organizes the deepest divergence in vertebrate evolution: reptiles and their descendants.
| Species | Genome | Total TE | DNA-TEs | Interpretation |
|---|---|---|---|---|
| Alligator | 2.18 Gb | 37.7% | 18.0% | Chaotic — active "cut & paste" |
| Turtle | 2.13 Gb | 15.0% | 3.3% | Stable — unchanged 200 Mya |
| Chicken | 1.05 Gb | 12.8% | 1.0% | Compact — ex-dinosaur, returned |
| Cattle | 2.67 Gb | 50.5% | 2.3% | Organized — BovB/L1 equilibrium |
The alligator — closest living relative of dinosaurs — harbors 18% DNA transposons, the most disruptive form of genomic chaos. The chicken — a direct dinosaur descendant — compressed its genome to half the mammalian size. The dinosaur's heir did not continue inflating. It returned to regulation.
As established in Section 1, the snake transferred BovB to mammals via horizontal transfer (Walsh et al. 2013; Ivancevic et al. 2018). BovB age in cattle (~22 Mya by divergence clock) is young relative to L1 (~43 Mya) — a recent, foreign arrival that was incorporated into the host's regulatory architecture. The promised consequence to the woman — "I will greatly multiply your pain in childbearing" (Genesis 3:16) — aligns with BovB's enrichment at reproductive and nefesh (physical vitality) genes: MHC, olfactory receptors, and reproductive loci are all BovB-enriched in ruminants.
The alligator, with 65 distinct TE families and 18% DNA transposons (the "cut and paste" class — the most disruptive form of transposon activity), represents genomic chaos. The turtle, unchanged for 200 million years with only 15% total TE and one dominant family (CR1 at 54%), represents ancient stability. The chicken — direct descendant of theropod dinosaurs that once produced the largest land animals in Earth's history — compressed its genome to 1.05 Gb (half the mammalian average), with only 12.8% TE content. The dinosaur's heir returned to regulation.
These are not abstractions. The alligator is the closest living relative of the organisms that dominated Earth for 165 million years. Its genome reads like an archaeological site: layer upon layer of independent transposon invasions, none organized, none in equilibrium. The cattle genome, by contrast, at 50.5% TE — more transposon content than the alligator — is structured: LINE elements (L1 and BovB) account for 28.3%, organized into two balanced systems. High content, but regulated. The organizing principle is not "how much" but "how managed."
11. Two Weeks, Two Genomes: The Reptilian Transition
The Hebrew Bible opens with two creation narratives that have long puzzled commentators. Traditional source criticism attributes the difference to separate documentary traditions, while harmonistic reading treats them as complementary perspectives on a single event. Neither approach, however, has attended to a fact that becomes striking once genomic data enter the conversation: the two accounts deploy different divine names, different verbs, different animal terminology — and these distinctions map, with unexpected precision, onto the difference between transposon-chaotic reptilian genomes and transposon-regulated avian and mammalian genomes. This section traces the mapping in detail.
Two Creation Accounts, Two Divine Names
Genesis 1 — what we will designate "Week One" — employs exclusively the name אלהים (Elohim). On Day 5 we read: "וַיִּבְרָא אֱלֹהִים אֶת הַתַּנִּינִם הַגְּדֹלִים... וְאֵת כָּל עוֹף כָּנָף" (Gen 1:21) — Elohim created the great tanninim … and every winged bird. On Day 6 the earth is commanded to bring forth "נֶפֶשׁ חַיָּה… בְּהֵמָה וָרֶמֶשׂ וְחַיְתוֹ אָרֶץ" (Gen 1:24) — living creatures … cattle, creeping things, and beasts of the earth. Three terms will prove critical: תַּנִּינִם הַגְּדֹלִים (great reptiles), עוֹף כָּנָף (winged bird, defined by its physical appendage), and חַיְתוֹ אָרֶץ (beasts of the earth, defined by raw matter). Throughout, the creative verb is בָּרָא (bara — to create ex nihilo or by radical origination), and the sole agent is אלהים.
Genesis 2 — "Week Two" — introduces a compound name that was absent from the first chapter: יהוה אלהים (YHWH Elohim). Here the animals are formed anew: "וַיִּצֶר יהוה אֱלֹהִים מִן הָאֲדָמָה כָּל חַיַּת הַשָּׂדֶה וְאֵת כָּל עוֹף הַשָּׁמָיִם" (Gen 2:19) — YHWH Elohim formed from the ground every beast of the field and every bird of the heavens. The verb has shifted from בָּרָא to יָצַר (yatsar — to form, to shape with directed intent, the verb used of a potter at the wheel). The animals have been renamed: the bird is no longer עוֹף כָּנָף (winged bird) but עוֹף הַשָּׁמַיִם (bird of the heavens); the beast is no longer חַיְתוֹ אָרֶץ (beast of the earth) but חַיַּת הַשָּׂדֶה (beast of the field). And the תַּנִּינִם הַגְּדֹלִים — the great reptiles — are entirely absent. They receive no mention in Week Two whatsoever. These are not stylistic variations; they are systematic lexical shifts, and they will prove to carry genomic weight.
The Linguistic Distinction
The morphological letter-group framework developed in earlier chapters allows us to unpack why the terminological shift matters. The word כָּנָף (wing) decomposes as כ (from the BKL group, denoting containment and instrumentality) + נ (from the AMTN group, denoting material substance) + ף (a Foundation letter). The composite meaning points to a physical appendage — the wing as a striking, covering, sheltering instrument. When Genesis 1 calls the bird עוֹף כָּנָף, it defines the creature by what it physically possesses: a mechanical apparatus for flight. The emphasis is on bodily equipment — structure without teleological orientation.
The word שָׁמַיִם (heavens), by contrast, decomposes as ש (a Foundation letter) + מ (AMTN) + י (YHW group, denoting directed will) + ם (AMTN), yielding a 25% Foundation-letter composition. שָׁמַיִם denotes not an organ but a domain — the heavenly realm to which the bird belongs. When Genesis 2 renames the bird עוֹף הַשָּׁמַיִם, it defines the creature not by mechanical equipment but by its place within a directed cosmic order. The bird is no longer characterized by what it has but by where it belongs.
A parallel shift governs the land animals. חַיְתוֹ אָרֶץ (beasts of the earth) ties the animal to אֶרֶץ — physical ground, raw materiality. חַיַּת הַשָּׂדֶה (beasts of the field) ties it instead to שָׂדֶה, a word cognate with שַׁדַּי (Shaddai, as in אֵל שַׁדַּי), evoking a bounded, cultivated, regulated domain rather than undifferentiated matter. The field is nature under governance; the earth is nature as substrate. Most telling of all is the fate of the תַּנִּינִם הַגְּדֹלִים: these "great reptiles" appear only in Week One, under אלהים alone, and never cross into the Week Two narrative. In the framework proposed here, they belong exclusively to the אלהים-only regime — the regime of multiplication without directed regulation.
The Genomic Mapping
The distinction between the two regimes maps onto comparative genomics with a coherence that merits careful attention.
Week One (אלהים only) — Transposon Chaos. The American alligator genome (Alligator mississippiensis) comprises approximately 37.7% transposable elements, with an exceptionally high 18% contributed by DNA transposons distributed across at least 65 distinct families (Green et al. 2014). This is genomic chaos in the technical sense: a proliferative, multi-family TE landscape with no single regulatory axis imposing order. Large dinosaurs — the creatures most naturally identified with the תַּנִּינִם הַגְּדֹלִים — are estimated to have carried genomes of 2–3 Gb on the basis of osteocyte cell-size proxies (Organ et al. 2007), consistent with the bloated, TE-laden architecture seen in extant crocodilians. In these genomes there is no BovB/L1 equilibrium, no L1HS-mediated management, no KRAB-ZFP silencing cascade. What there is, is multiplication — raw, undirected, prolific insertion of mobile elements. This is precisely the regime of אלהים, the divine name associated throughout Genesis 1 with the command פְּרוּ וּרְבוּ (be fruitful and multiply) — creation as quantitative expansion governed by natural law, without the superimposed layer of covenantal direction.
Week Two (יהוה אלהים) — Regulated Genomes. Modern birds, descended from theropod dinosaurs or (on an alternative reading) separately formed, carry strikingly compact genomes: the chicken genome is approximately 1.05 Gb, with only 12.8% TE content (International Chicken Genome Sequencing Consortium 2004). The contrast with the alligator is enormous — a roughly threefold reduction in genome size and a near-threefold reduction in TE proportion. Among mammals, ruminants maintain a precisely calibrated BovB/L1 equilibrium in which the two retrotransposon lineages constrain one another's copy number (as discussed in Section 8.3). Across the mammalian clade more broadly, piRNA pathways and KRAB-zinc-finger proteins provide multi-layered TE surveillance, actively silencing insertions that threaten genomic integrity. These are genomes under governance — not merely multiplying, but regulated. In the two-week framework, the addition of the name יהוה to אלהים signifies precisely this superimposition: directed regulation layered upon automatic multiplication. The verb יָצַר (formed) captures the difference — the potter does not create matter but shapes what already exists toward an intended form.
The Snake Recognizes Only One Name
A detail in Genesis 3 acquires new significance in this framework. When the serpent addresses Eve, it says: "אַף כִּי אָמַר אֱלֹהִים לֹא תֹאכְלוּ" — Did Elohim indeed say you shall not eat? (Gen 3:1). The serpent speaks only the name אלהים. It never utters יהוה. In the traditional commentarial literature this is often noted as evidence of the serpent's spiritual limitation — the creature cannot access the covenantal dimension represented by the Tetragrammaton. Within the morphological framework proposed here, the observation acquires a genomic correlate. As demonstrated in Section 8.2, BovB content in snakes stands at approximately 0.01% — a vanishingly small remnant indicating that snakes served as the original horizontal donors of BovB to mammalian genomes. The serpent gave everything and retained nothing. It operates in the אלהים-only regime: pure multiplication, pure transmission, without the internal regulatory architecture that characterizes the יהוה אלהים domain. The snake neither possesses BovB/L1 equilibrium nor deploys KRAB-ZFP silencing. It is, genomically, a creature of Week One that persists into the present — and its speech, restricted to a single divine name, encodes exactly this status.
Time in Week One
A frequently raised objection concerns chronology: how can the "days" of Genesis 1 accommodate the Mesozoic Era, a span of roughly 186 million years during which the great reptiles dominated? The text itself provides a remarkable answer. The sun, moon, and stars — the instruments of measurable time — are not created until Day 4: "יְהִי מְאֹרֹת בִּרְקִיעַ הַשָּׁמַיִם… לְאֹתֹת וּלְמוֹעֲדִים וּלְיָמִים וְשָׁנִים" — Let there be luminaries in the firmament of the heavens … for signs, and for seasons, and for days, and for years (Gen 1:14). Prior to Day 4, there exists no solar metric by which a "day" can be defined. The text explicitly states that calendrical time — days, seasons, years — begins only with the appearance of the luminaries. It follows that the "days" preceding Day 4 are not solar days; they are epochs of indeterminate duration operating under the אלהים regime, in which time as a measured quantity does not yet exist. The great reptiles of Day 5 — the תַּנִּינִם הַגְּדֹלִים — inhabit this pre-calendrical expanse. The Mesozoic fits naturally into the framework, not by allegorizing the text but by reading it on its own terms: measurable time begins when the text says it begins, and not before.
The Flood as Regime Boundary
The Flood narrative functions within this framework as the boundary event between the two genomic regimes. What enters the ark is selected — chosen, preserved, carried into the post-diluvian world that operates under the יהוה אלהים dispensation. What remains outside the ark belongs to Week One — the אלהים-only regime — and perishes. The ark thus represents a genomic bottleneck of the most extreme kind: eight human survivors, a minimum viable population of each animal kind, and a catastrophic collapse in genetic diversity. As argued in earlier sections of this chapter, such a bottleneck would devastate piRNA diversity, degrading the regulatory machinery that suppresses transposon activity and triggering the observed 35.6-fold reduction in human lifespan from the antediluvian to postdiluvian periods.
It is not necessary to identify the Flood with the K-Pg extinction event of approximately 66 million years ago, although the structural parallel is striking: in both cases, a catastrophic boundary separates a TE-chaotic regime (Mesozoic reptiles; antediluvian world) from a TE-regulated regime (Cenozoic birds and mammals; postdiluvian humanity). Both events function as regime transitions — from chaos to regulation, from unbounded multiplication to managed complexity. The two-week framework does not require harmonizing biblical and geological chronologies; it requires only that we recognize the structural identity of the transition.
What This Does Not Claim
Intellectual honesty demands explicit statement of the framework's boundaries. We do not claim that paleontology is mistaken about bird–dinosaur descent. Whether modern birds evolved from theropod dinosaurs through a gradual process of genome streamlining or were separately formed by a divine act, the genomic transition is empirically identical: from a TE content exceeding 35% (reptilian architecture) to a TE content of approximately 13% (avian architecture), accompanied by a threefold reduction in genome size. The two-week framework offers a structural reading of Genesis that maps onto this genomic data; it does not require the reader to reject evolutionary timelines, nor does it depend on any particular stance regarding common descent. The mechanism is the same regardless of one's metaphysical commitments: regulation replaced chaos, directed management superseded undirected multiplication, and the genomic signature of that transition is written into every bird genome sequenced to date.
What the framework does claim is that the terminological precision of Genesis — the shift from כָּנָף to שָׁמַיִם, from אָרֶץ to שָׂדֶה, from בָּרָא to יָצַר, from אלהים alone to יהוה אלהים together — encodes a distinction that is not merely theological but genomic. The two divine names correspond to two modes of biological organization: multiplication without regulation, and multiplication under regulation. The great reptiles belong to the first; the birds of the heavens and beasts of the field belong to the second. And the serpent, knowing only one name, belongs irreversibly to the regime it never left.
Summary. Genesis 1 (אלהים alone) describes multiplication without directed regulation — matching the reptilian TE chaos exemplified by the alligator genome (37.7% TE, 65 transposon families). Genesis 2 (יהוה אלהים) introduces directed regulation — matching compact avian genomes (1.05 Gb, 12.8% TE) and the managed transposon landscapes of mammals. The terminological shifts are systematic: עוֹף כָּנָף (physical wing) gives way to עוֹף הַשָּׁמַיִם (heavenly domain); חַיְתוֹ אָרֶץ (beasts of the earth) gives way to חַיַּת הַשָּׂדֶה (beasts of the field). The תַּנִּינִם הַגְּדֹלִים appear only under אלהים and vanish from the narrative entirely. The serpent, confined to the name אלהים, operates genomically in the regime of pure transmission without regulation (BovB ≈ 0.01%). The Flood functions as the regime boundary, and the pre-Day-4 absence of solar time accommodates epochs of indeterminate duration. The framework maps text onto data without requiring acceptance or rejection of evolutionary chronology.
12. The Matter/Spirit Equation
The data presented thus far — BovB/L1 ratios, L1HS activation, TE depletion, reptilian baselines — describe a physical system. But the Torah encodes the same distinctions in its morphology. The Foundation percentage (F%) analysis, validated across 98,122 word pairs with 87.8% predictive accuracy, assigns each letter to a functional group. The resulting gradient maps directly onto the genomic data.
The Foundation percentage (F%) assigns each Hebrew letter to one of four functional groups. Applied to names encoding the physical-spiritual polarity:
| F% | Category | Examples | Genomic parallel |
|---|---|---|---|
| 0% | Pure regulation | יהוה, אלהים | L1HS management system |
| 25% | Activation | נשמה, אד-ני | L1HS itself — the switch |
| 40% | Elevated human | ישראל | Regulated L1HS + Torah |
| 50% | Base human | יעקב | L1HS active, pre-elevation |
| 67% | Matter-heavy | עשו, עפר | Ape phenotype: L1HS off |
| 75% | Excess matter | שעיר | KRTAP overexpression |
| 100% | Pure matter | שער (hair) | Keratin — most BovB-rich tissue |
The name change from יעקב (50%) to ישראל (40%) encodes a regulatory upgrade — less matter, more control. The word אדמה (earth, 25% F) gains one letter to become אדם (human) — and that letter is ה, a YHW differentiation letter. In the morphological system we have documented across 98,122 word pairs, YHW letters are the precise mechanism by which roots differentiate into distinct meanings.
The genome that all mammals share is the אדמה. The layer that makes one species human is the ה.
13. Rapid Diversification: The BovB Engine
The preceding sections establish regulatory architecture as the organizing principle of existing species. A natural question follows: can the same mechanism explain the origin of species diversity?
The feasibility of rapid diversification from a limited ancestral pool:
Precedent. Cichlid fish in Lake Victoria: ~500 species from 1 ancestor in ~15,000 years (Seehausen 2006). Peer-reviewed, non-controversial.
Mechanism. BovB at different loci → different phenotypes. BovB at KRTAP → keratin horns (Bovidae). BovB at SHH → fangs (Moschidae). Same transposon, different target, different species. Insertion sites: 94.5% shared cow↔sheep (±10 kb), 74% cow↔deer, 51.3% sheep↔deer — phylogenetic gradient from common origin.
Rate. BovB is still active in cattle: 7,325 copies at <2% divergence from consensus, indicating ongoing transposition. Sheep show 4,854 young copies; deer only 427 — a gradient of decreasing BovB activity that mirrors the BovB/L1 ratio itself. With approximately 1 new insertion per generation across ~1,667 generations in 5,000 years, the ruminant lineage would have experienced ~133,000 insertion opportunities. To produce 200 species from 10 ancestral types requires a "success rate" of 0.15%.
Karyotype evidence. The muntjac provides living proof of TE-driven chromosomal change: Muntiacus muntjak has 2n=6 (the lowest chromosome number of any mammal), while Reeves' muntjac has 2n=46 — within a single genus. These are not ancient divergences. They demonstrate that transposon-mediated chromosome fusion and fission can radically restructure a genome in evolutionary short timescales. Fedoroff (2012, Science) and Chuong et al. (2017, Nature Reviews Genetics) have independently argued that transposable elements serve as speciation engines, providing the raw material for reproductive isolation and regulatory innovation.
14. Three Levels of Regulation
The data now span ruminants, primates, reptiles, growth genes, and speciation mechanisms. A pattern emerges that is not visible from any single dataset but becomes clear when they are read together: biological regulation operates in hierarchical levels, each building on the previous.
The data reveal three hierarchical levels of biological regulation, each building on the previous:
Level 1: בהמה — Automatic regulation. The BovB/L1 equilibrium in ruminants operates without cognition. The ratio maintains itself through molecular mechanisms — selection, methylation, insertion preference. The Hebrew word בהמה may be read as ב-ה-מ-ה: "in her are the forces." This is regulation as homeostasis. The sheep at 1.00 is its purest expression.
Level 2: אדם — Conscious regulation. L1HS is active in human neurons — specifically in the hippocampus, the seat of learning and memory. Coufal et al. (2009) demonstrated L1 retrotransposition in neural progenitor cells. Each new insertion alters gene expression in that neuron, creating unique regulatory states. This is not homeostasis. It is adaptation. Learning. Choice. The regulation is no longer automatic — it is managed by consciousness.
Level 3: תורה — Directed regulation. If L1HS provides the mechanism for neuroplasticity, and neuroplasticity enables learning, then a system of directed learning — structured behavioral practice — constitutes a third regulatory layer. The Torah (from the root הוראה, "instruction") provides 613 behavioral protocols. Each behavioral change propagates through well-documented pathways: behavior → neural activity → epigenetic modification → gene expression change. The Torah does not describe regulation. The Torah is regulation — a firmware layer for the conscious regulatory system that L1HS enables.
This hierarchy resolves a question the data raise but cannot answer on their own: why is L1HS active in humans and not in apes? The molecular answer (stronger APOBEC3B/PIWIL2 defense) explains how. The regulatory hierarchy suggests why: L1HS requires a management system. In the animal, BovB/L1 is self-managing. In the human, L1HS requires conscious management. Without a system of directed practice — without "instructions" — the switch is dangerous (Rett syndrome) rather than productive.
The molecular pathway from behavior to gene expression is well-documented: behavioral engagement → neural firing → immediate early gene activation (cFos, BDNF) → histone acetylation → chromatin remodeling → L1 demethylation → new L1 insertion → altered gene expression in that neuron. Maze et al. (2011) demonstrated this pathway directly: cocaine exposure activated L1 retrotransposition in the nucleus accumbens, altering reward circuitry. The pathway is real, published, and not specific to pathology — it operates whenever sustained neural engagement produces epigenetic change.
Torah study, in this framework, is structured neural engagement across all behavioral domains. The 613 commandments span diet (kashrut), sleep (Shema before bed), speech (laws of lashon hara), work (Shabbat rest), reproduction (family purity), agriculture (shmita), and justice (courts). No behavioral domain is unregulated. Each behavioral protocol, practiced consistently, engages specific neural circuits, produces specific epigenetic changes, and — if the L1HS mechanism operates as documented — produces specific regulatory states in specific neurons. The Torah is not a description of regulation. It is a behavioral protocol that, through documented molecular pathways, produces regulation.
15. Genesis 3: The Regulatory Event
The Torah's account of the Garden of Eden, read through the regulatory framework established in this chapter, describes a specific genomic transition: from automatic regulation to conscious regulation, triggered by the introduction of a foreign genetic element.
The Agent: Pure Matter
The Foundation percentage of the narrative's key agents is striking:
| Word | Letters | F% | Category |
|---|---|---|---|
| נחש (serpent) | נ(A)+ח(F)+ש(F) | 67% | Matter-heavy |
| זרע (seed) | ז(F)+ר(F)+ע(F) | 100% | Pure matter |
The word זרע — seed, offspring, genetic material — is 100% Foundation. Every letter is a content letter. In a language where the divine names contain 0% Foundation, the word for the serpent's genetic contribution is pure physical substance.
By contrast, the post-event vocabulary shifts dramatically:
| Word | F% | Category |
|---|---|---|
| דעת (knowledge) | 33% | Regulatory |
| מות (death) | 0% | Pure regulatory |
| איבה (enmity) | 0% | Pure regulatory |
| כתנת (garment) | 0% | Pure regulatory |
The "before" words (טוב, חיים, ערום) average 42.9% Foundation. The "after" words (דעת, מות, איבה, כתנת, עור) average 33.3% — a 9.5 percentage point drop. The narrative moves toward greater regulatory content, not less. "Knowledge of good and evil" is, morphologically, an increase in regulatory capacity.
The Guardians: TE-Depleted Defense Genes
Genesis 3:24 describes what was placed to guard the path back to the original state:
"ויגרש את האדם וישכן מקדם לגן עדן את הכרבים ואת להט החרב המתהפכת לשמר את דרך עץ החיים"
"He placed the cherubim and the flaming sword that turns, to guard the way to the tree of life."
The genome contains a defense system against L1 retrotransposition — the very element whose activation distinguishes humans from apes. We measured TE density at the genes encoding this defense:
| Gene | Function | TE × genome | Status |
|---|---|---|---|
| PIWIL1 | piRNA pathway (L1 silencing) | ×0.56 | Protected |
| DNMT3A | De novo methylation | ×0.72 | Protected |
| PIWIL2 | piRNA pathway | ×0.89 | Mildly depleted |
| DNMT3B | De novo methylation | ×0.86 | Mildly depleted |
| APOBEC3B | L1 RNA destruction | ×1.16 | Not depleted |
| PIWIL4 | piRNA pathway | ×1.88 | L1-invaded |
PIWIL1, the primary piRNA defense gene, is TE-depleted at ×0.56 — approaching the protection level of IGF2 (×0.37), the most guarded gene in the genome. DNMT3A, the de novo methyltransferase that writes the methyl marks silencing L1, is protected at ×0.72. These are the genome's "cherubim" — static guardians, TE-depleted, maintaining the barrier between active L1 and the autoregulated state.
APOBEC3B operates differently: it is an active enzyme that deaminates L1 RNA, destroying it chemically. It is not TE-depleted (×1.16) because it does not need passive protection — it is itself a weapon. A "flaming sword."
The word מתהפכת (turning, revolving) adds precision: APOBEC3B's defense is not permanent. It resets each generation when methylation is reprogrammed in the germline. Every generation, the sword "turns" — the defense must be re-established. Baduel et al. (2025) documented this generational reset in mammals.
And PIWIL4 — a piRNA gene tasked with silencing L1 — has been invaded by L1 at ×1.88. The serpent is inside the guardian. The arms race described by Jacobs et al. (2014) is not a metaphor applied to Genesis. It is a measured reality to which Genesis maps.
Two Events, Two Elements
The narrative distinguishes two events:
1. The serpent's contribution — "between your seed (זרע) and her seed" (Genesis 3:15). BovB: a horizontally transferred reptilian retrotransposon, documented by Walsh et al. (2013). The "seed of the serpent" is, genomically, a measured reality.
2. The tree's consequence — "knowing good and evil" (דעת טוב ורע). The acquisition of regulatory capacity: the ability to distinguish, to manage, to choose. In the genome, this maps to the L1HS management system — APOBEC3B, PIWIL2, the methylation apparatus — that permits controlled L1 activity in the human brain.
Before the event: automatic regulation. The "tree of life" — עץ החיים — represents a state where BovB is absent and L1 does not require conscious management. This is Level 1: בהמה.
After the event: conscious regulation required. BovB is now present in the genome (the serpent's seed). L1 can no longer be left on autopilot. The human must develop and maintain a management system — or suffer the consequences (Rett syndrome: MeCP2 failure → uncontrolled L1 → neurological collapse).
"Garments of skin" — כתנות עור (Genesis 3:21) — are the first thing provided after the regulatory transition. Skin is the tissue with the highest BovB enrichment in the mammalian body: KRTAP at 22.5%. The garment is not a metaphor for modesty. It is the first BovB-rich tissue — the physical mark of the new genomic state.
The Fig Leaf: First Covering
Genesis 3:7 records the first response to the acquisition of knowledge: "ויתפרו עלה תאנה" — "they sewed fig leaves." The choice of fig is not incidental.
The fig (תאנה: ת-א-נ-ה) is 0% Foundation — pure regulatory letters, no physical content. It shares this property with the divine names יהוה and אלהים. The wasp (צרעה: צ-ר-ע-ה) that pollinates it is 75% Foundation — matter-heavy, and it contains within its letters the word רע (evil, 100% Foundation).
The fig cannot reproduce without the wasp. The wasp dies inside the fig. This is not metaphor — it is documented obligate mutualism. The fig wasp (Blastophaga psenes) enters the fig, pollinates it, and dies within. Without this death, the fig produces no fruit. The "evil" (רע, 100% matter) must enter the "pure" (תאנה, 0% matter) and be consumed for life to continue.
The first covering is regulatory (fig leaf, 0% F). The second covering, provided by God, is physical: כתנות עור — "garments of skin" (Genesis 3:21). Skin is the tissue with the highest BovB enrichment: KRTAP at 22.5%. The narrative moves from regulatory response to physical reality — from 0% Foundation to the most BovB-rich tissue in the mammalian body.
The Virus That Became Motherhood
The pattern of foreign elements becoming essential for life extends beyond transposons. Syncytin-1, the protein essential for formation of the syncytiotrophoblast layer of the human placenta, is encoded by the envelope gene of an endogenous retrovirus (HERV-W). Mi et al. (2000, Nature 403:785) demonstrated that syncytin mediates cell-cell fusion in the placenta. Without it, no placenta forms; without placenta, no mammalian reproduction.
Syncytin has been independently captured at least six times across mammalian lineages (Lavialle et al. 2013) — different retroviruses, same essential function, convergent domestication. A "parasite" became the sine qua non of motherhood.
Genesis 3:20: "ויקרא האדם שם אשתו חוה כי היא היתה אם כל חי" — "The man named his wife Eve, because she was the mother of all living." To be the mother of all living, in mammals, requires a placenta. To have a placenta requires a retrovirus. The naming occurs immediately after the curses, as if acknowledging: the new reproductive reality — dependent on a viral gene — is now the path forward.
Syncytin is not an isolated case. Approximately 5–8% of the human genome consists of endogenous retroviral sequences — three to five times more viral DNA than protein-coding DNA. Among the documented domestications: HERV-K is active from the 8-cell embryo stage and its Rec protein provides antiviral defense (Grow et al. 2015, Nature). MER41 elements serve as interferon-inducible enhancers; their CRISPR deletion collapses the innate immune response (Chuong, Elde & Feschotte 2016, Science). Ninety percent of endogenous retroviral remnants are solo LTRs functioning as promoters and enhancers, with 64% of LTR-driven transcripts expressed in reproductive tissues. The human genome is not merely "contaminated" by viruses. It is operated by them.
The Lifespan Collapse
The Torah records a precise decline in human lifespan across generations:
| Generation | Name | Lifespan | Phase |
|---|---|---|---|
| 1 | Adam | 930 | Pre-flood plateau |
| 7 | Methuselah | 969 | Pre-flood plateau |
| 10 | Noah | 950 | Last pre-flood |
| 11 | Shem | 600 | Post-flood collapse |
| 14 | Eber | 464 | |
| 16 | Peleg | 239 | |
| 20 | Abraham | 175 | |
| 22 | Jacob | 147 | |
| 26 | Moses | 120 | Asymptotic limit |
Fitting an exponential decay model reveals two distinct phases: - \1 decay constant k = 0.0034 (near-flat plateau at ~930 years) - \1 decay constant k = 0.1207 (\1 than pre-flood)
The collapse is not gradual. It is abrupt — beginning immediately after the flood bottleneck, when the human population contracted to eight individuals.
De Cecco et al. (2019, Nature) demonstrated that L1 retrotransposons activate during aging in somatic cells, triggering the cGAS-STING innate immune pathway and producing chronic inflammation — the molecular signature of aging. L1 activation is not a consequence of aging; it is a driver of aging.
A population bottleneck of eight individuals would catastrophically reduce the diversity of piRNA sequences, KRAB-ZFP variants, and methylation patterns that collectively silence transposable elements. Each subsequent generation, with reduced silencing diversity, would experience greater TE derepression — cumulative, compounding, exponential.
Genesis 6:3: "והיו ימיו מאה ועשרים שנה" — "His days shall be one hundred and twenty years." The exponential decay curve asymptotes at approximately 120 — the limit specified before the flood occurred. Moses lived exactly 120 years (Deuteronomy 34:7).
16. The Universal Pattern: Foreign Becomes Essential
Three independent biological systems demonstrate the same principle:
| System | Foreign element | Host | Result | Torah reference |
|---|---|---|---|---|
| Ruminant genome | BovB (snake) | L1 (mammal) | BovB/L1 equilibrium → kashrut | Genesis 3:14-15 |
| Human placenta | HERV-W (retrovirus) | Uterus | Syncytin → reproduction | Genesis 3:20 |
| Fig reproduction | Wasp (צרעה, 75%F) | Fig (תאנה, 0%F) | Obligate mutualism → fruit | Genesis 3:7 |
| Human immunity | MER41 (retroviral LTR) | Interferon genes | Enhancer → immune response | — |
The Fig and the Wasp: A Detailed Case
The fig-wasp mutualism deserves closer examination because its mechanism is the most physically dramatic of the five instances.
The unripe fig is toxic — sealed, filled with tannins and latex, inedible. A single female wasp of the species-matched partner (approximately 500 Ficus species each paired with one specific wasp species — no substitutions possible) enters through the ostiole, breaking her wings and antennae in the process. She pollinates the internal flowers, lays her eggs, and dies inside the fig.
The fig's ficin protease then completely dissolves the wasp's body — she is absorbed as nutrition for the developing fruit. Pollination triggers an ethylene cascade: tannins degrade (bitter → neutral), sugars accumulate (acid → sweet), the fruit softens and changes color. The result is one of nature's sweetest fruits, a keystone species feeding over 1,200 vertebrate species.
The fig system actually requires two trees: the caprifig (male, which houses the wasp colony — many wasps, no edible fruit) and the fig proper (female, which receives one wasp, produces fruit). The caprifig is the reservoir; the fig is the recipient. Many → one → sacrifice → life.
This is irreducible co-dependence. The fig cannot reproduce without the wasp. The wasp cannot reproduce without the fig. Neither can exist without the other already existing. The 500 species-specific pairs represent 500 instances of simultaneous co-creation — a system that cannot be built incrementally. Seventy to ninety million years of strict cospeciation confirm that these pairs have maintained their lock-and-key specificity across geological time.
The Torah places the fig leaf as the first response to the acquisition of knowledge (Genesis 3:7). The cherubim (כרובים — always plural, always paired) guard the tree of life (Genesis 3:24). The word כרוב (cherub) is 50% BKL — relation letters. Cherubim are, morphologically, about relationship: two entities, facing each other (Exodus 25:20: "ופניהם איש אל אחיו"), neither functional alone. Two fig trees. Two guardians. Two systems — defense and life — that require each other.
In each case: 1. A foreign organism contributes genetic material or physical presence to a host. 2. The contribution is initially "parasitic" or destructive. 3. Through regulatory domestication, the foreign element becomes \1. 4. The Torah references all three within the same narrative (Genesis 3).
The F% encoding is consistent:
| Element | F% | Role |
|---|---|---|
| זרע (seed) | 100% | Foreign genetic material |
| רע (evil) | 100% | Unregulated matter |
| צרעה (wasp) | 75% | Foreign agent (contains רע) |
| נחש (serpent) | 67% | Foreign agent |
| תאנה (fig) | 0% | Host — pure regulation |
| איבה (enmity) | 0% | Regulatory tension |
| חיים (life) | 25%F, 50% YHW | Differentiation = life |
The word רע does not encode moral evil. It encodes unregulated matter — 100% Foundation, zero control letters. Evil, in this morphological system, is physical substance without regulatory management. Fire without a furnace. L1 without methylation. BovB without L1 counterbalance.
Regulated רע = life. The wasp dies in the fig, and the fig bears fruit. BovB enters the mammalian genome, and the altar animal achieves equilibrium. A retrovirus integrates into the human genome, and the placenta forms. In every case, what was foreign, destructive, or "evil" becomes — through regulation — the mechanism of life itself.
Figure 4. Biblical lifespan decay: ×35.6 post-flood.
17. Three Additional Convergences
The Ox and the Donkey: Dual and Mono Systems
"You shall not plow with an ox and a donkey together" (Deuteronomy 22:10).
| Animal | BovB% | L1% | BovB/L1 | System |
|---|---|---|---|---|
| Ox (שור) | 13.33% | 12.95% | 0.97 | Dual (BovB + L1) |
| Donkey (חמור) | 0.00% | 12.38% | 0.00 | Mono (L1 only) |
The ox operates a dual regulatory system — two balanced retrotransposon families in near-equilibrium. The donkey operates a mono system — L1 only, no BovB. The prohibition against yoking them together separates two fundamentally different genomic architectures. One is regulated by tension between two forces; the other has no such tension. Combining them in a single task conflates two systems that operate on incompatible principles — like connecting circuits running at different voltages.
Chametz and Matzah: Inflated and Compressed Genomes
The five grains that can become chametz (leaven) — wheat, barley, spelt, rye, oats — have among the most inflated genomes in the plant kingdom. Wheat (Triticum aestivum) has a 17 GB genome, 85% repetitive sequence, hexaploid. Rice (Oryza sativa), which cannot become chametz, has a 430 MB genome — ×40 smaller, 35% repetitive.
| Grain | F% | Genome | Repeats | Status |
|---|---|---|---|---|
| חמץ (leaven) | 67% | — | — | Forbidden on Pesach |
| מצה (matzah) | 33% | — | — | Required on Pesach |
| חטה (wheat) | 67% | 17 GB | 85% | Can become chametz |
| אורז (rice) | 50% | 430 MB | 35% | Cannot become chametz |
Chametz (חמץ, 67% Foundation) shares its root with חמצון — oxidation. Leavening is a biological process: yeast fermentation inflates dough, just as transposon amplification inflates genomes. Matzah (מצה, 33% Foundation, 33% YHW) is the compressed form — dough before inflation, genome before TE expansion.
Pesach mandates the annual transition from chametz to matzah, accompanied by the Passover lamb — the animal at BovB/L1 = 1.00. The Pesach protocol requires eating the compressed genome (matzah) together with the equilibrium genome (lamb), while avoiding the inflated genome (chametz). Failure to comply carries the penalty of כרת (Exodus 12:15) — the same penalty as failing to offer the Passover sacrifice itself.
The Vine and the Wine: Matter Becomes Spirit
Genesis 9:20: "ויחל נח איש האדמה ויטע כרם" — "Noah, man of the earth, began and planted a vineyard." The first act of the post-flood world is planting a vine.
| Word | F% | YHW% | Interpretation |
|---|---|---|---|
| גפן (vine) | 67% | 0% | Raw matter — the plant |
| יין (wine) | 0% | 67% | Pure spirit — the product |
| שכר (intoxicant) | 67% | 0% | Matter — unregulated consumption |
The vine (גפן, 67% Foundation) is matter. Wine (יין, 0% Foundation, 67% YHW) is spirit — morphologically identical to the divine names. The transformation of grape to wine — fermentation, a biological process of controlled decomposition — converts pure matter into pure spirit. The same raw material, regulated differently, produces opposite F% signatures.
But שכר (intoxicant, 67% Foundation) — uncontrolled consumption of the same substance — returns to matter. The product is the same; the regulation differs. Wine for kiddush (sanctification) = 0% F. Wine for drunkenness (שכר) = 67% F. Regulation determines whether the same substance serves spirit or matter.
Noah — נח, 50% Foundation — is precisely balanced, as is the name itself. His first act in the new world is to create the substance that, properly regulated, embodies the principle this entire chapter describes: foreign or raw material, managed through regulation, becomes sacred.
The Nazir: Blocking the Pathway
The Nazir vow (Numbers 6) provides the inverse test. The Nazir must avoid every part of the vine — not only wine (the product) but grapes, seeds (חרצן), husks (זג), and vinegar: "מחרצנים ועד זג לא יאכל" — "from seed to husk he shall not eat." The entire biological unit is prohibited.
The F% of the vine's components reveals a gradient:
| Part | F% | Status |
|---|---|---|
| זג (husk) | 100% | Pure matter |
| חרצן (seed) | 75% | Mostly matter |
| ענב (grape) | 33% | Balanced |
| יין (wine) | 0% | Pure spirit |
The vine pathway runs from 100% matter (husk) to 0% matter (wine) — a complete transformation. The Nazir blocks this entire pathway. And the result? "תער לא יעבור על ראשו... גדל פרע שער ראשו" — "no razor shall pass over his head; he shall grow the hair of his head wild."
שער (hair) = 100% Foundation. פרע (wild growth) = 100% Foundation. Both equal to זג (husk) — 100% Foundation. Hair is the grape's husk, linguistically and morphologically: pure matter.
When the vine's matter-to-spirit pathway is blocked, matter accumulates — as hair. This is the same principle observed in primates: when L1HS (the regulatory switch) is off, the physical phenotype dominates — excessive hair, enlarged musculature, prominent jaw. The Nazir, by voluntarily blocking a regulatory pathway, temporarily enters a matter-dominant state. His hair — שער, KRTAP, 22.5% BovB — grows unchecked.
Remarkably, resveratrol — the polyphenol concentrated in grape skin — promotes hair growth in published studies (Zhang et al. 2021; Juchaux et al. 2020), while alcohol consumption is associated with hair loss (OR 1.4–1.9 for androgenic alopecia; Yang et al. 2024). The grape's raw material grows hair; the grape's transformed product (wine) inhibits it. The Nazir avoids the transformation and retains the raw effect.
18. What This Does Not Prove
Every finding in this chapter has an alternative explanation. Intellectual honesty requires stating them.
This chapter does not prove that the Torah's creation account is literally accurate. It does not prove that evolution did not occur. It does not prove that humans are not descended from earlier primates.
L1HS and L1Pt (the chimpanzee-specific variant) are phylogenetically sister lineages, both derived from L1PA2 (Lee et al. 2007). The data are consistent with a common ancestor — which, in the Torah's framework, is Adam. Whether Adam is a historical individual, a population, or a metaphor for the genomic template from which all primates diverged is a question the data constrain but do not resolve.
What the data clearly show:
1. L1 forms chronological layers — a base genome shared by all mammals, topped by species-specific regulatory elements. The creation order in Genesis 1 (land animals, then humans from the same earth) maps onto this architecture.
2. The human-ape difference is regulatory, not structural. Same genes, same TEs, same base genome (95% shared). One switch: L1HS.
3. The switch is epigenetic — methylation-based, reversible, reset every generation. It is not a permanent mutation but an ongoing regulatory commitment.
4. L1HS correlates with cognition and lifespan, not disease. The "parasite" narrative fails empirically.
5. The Torah's content matches the genomics. The snake transferred BovB. The sheep is at equilibrium. The Red Heifer is a reference standard. Hair is pure matter. The name change from Jacob to Israel is a regulatory upgrade. The breath of life is the activation layer. The giants are regulatory breaches.
There are honest caveats. The BovB/L1 correlation with Torah classification, while statistically significant, could in principle be coincidental — dietary laws might track some other biological feature (digestive anatomy, for instance) that happens to correlate with transposon content. The molecular clock assumptions underlying age estimates are contested, and TE divergence rates may vary across lineages. The F% analysis, while validated across 98,122 word pairs, could reflect a property of Hebrew phonology rather than an intentional encoding.
But the force of the argument lies not in any single correlation. It lies in three independent confirmations:
1. Language: The morphological system of Biblical Hebrew (Z=152.16, p<0.0001) cannot be replicated by any known human language, modern or ancient, including closely related Aramaic (Z=0.39, not significant).
2. Structure: The divine names function as morphological state indicators with predictive power (87.8% meaning prediction in 5-fold cross-validation on 98,122 pairs).
3. Content: The biological descriptions — snake as BovB donor, sheep at unity, red as diagnostic color, dust as shared genome, breath as activation layer, hair as pure matter, giants as regulatory breaches — align with measured genomic data.
The probability that all three are independently coincidental is the product of their individual probabilities. If each has even a generous 10% chance of being accidental, the combined probability is 0.1%. If the individual probabilities are closer to their measured p-values, the combined probability approaches zero.
None of this constitutes proof of divine authorship. All of it constitutes evidence that the Torah's biological content is not mythology — it is a description of regulatory architecture, written in a language whose own morphology demonstrates the same regulatory principles it describes.
The chapter began with the sheep at BovB/L1 = 1.00 and ends with the cherubim guarding the tree of life. Between them lies a single principle: regulation produces function; its absence produces either silence or chaos. This principle operates at the scale of transposons (BovB/L1 in ruminants), chromosomes (L1HS in primates), tissues (KRTAP in skin), neural circuits (L1 in hippocampal neurons), behavior (Torah as regulatory protocol), and narrative (Genesis 3 as the story of a regulatory transition). One architecture, expressed at every scale the data permit us to examine.
"וייצר יהוה אלהים את האדם עפר מן האדמה ויפח באפיו נשמת חיים ויהי האדם לנפש חיה"
(Genesis 2:7)
He formed the human from dust of the earth and breathed into his nostrils the breath of life, and the human became a living soul.
The dust — עפר, 67% Foundation — is the shared mammalian genome. The breath — נשמה, 25% Foundation — is the L1HS activation. The serpent's seed — זרע, 100% Foundation — is the BovB that entered. The evil — רע, 100% Foundation — is matter unregulated. The knowledge — דעת, 33% Foundation — is the regulatory capacity that followed. The fig leaf — תאנה, 0% Foundation — is the first regulatory response. The guardians — כרובים, PIWIL1 at ×0.56 — still stand watch. The lifespan — 950 to 120, ×35.6 after the bottleneck — is L1 running unsilenced. The virus — syncytin, HERV-W — became the placenta, and the mother of all living. The instructions — תורה, from הוראה — are the operating system.
One architecture. Letters to DNA. Serpent to guardian. Wasp to fig. Vine to wine. Evil to life. Instruction to regulation.
Summary
This chapter presents a single thesis: regulation, not complexity, is the organizing principle of the genome — and this principle aligns with the Torah's biological descriptions.
The evidence spans four domains:
Ruminants. The BovB/L1 ratio — measuring the balance between snake-derived and endogenous retrotransposons — distinguishes altar animals (0.94–1.00) from kosher non-altar (0.63) from forbidden (0–0.001). The same ratio determines cranial morphology: KRTAP enrichment → keratin horns; SHH enrichment → fangs. Zero exceptions across six ruminant families. The snake transferred BovB (Walsh 2013); the altar animals domesticated it.
Primates. L1HS — the only actively transposing retrotransposon in any primate genome — exists at 1,536 copies in humans and 69–78 remnants in apes. The genes are identical (FOXP2, ASPM, KRTAP: same TE density). The difference is one epigenetic switch: methylation-based, reversible, reset every generation. Neanderthals (96% of human L1HS sites absent) represent the intermediate: large brain, inactive switch.
Development. Growth genes (IGF2 ×0.37, GH1 ×0.66) are systematically guarded from transposon insertion. Breaching this protection produces gigantism — a phenotype the Torah describes as rare, post-flood, and individually named.
Language. The Foundation percentage (F%) maps the same polarity: שער (hair) = 100% matter; יהוה = 0% matter. The word אדם (human) differs from אדמה (earth) by one differentiation letter: ה. The genome encodes the same distinction — shared base (95% L1M), unique activation (L1HS).
Three levels of regulation emerge: automatic (בהמה: BovB/L1 equilibrium), conscious (אדם: L1HS in neurons), and directed (תורה: behavioral protocols producing epigenetic change). The Torah does not describe this architecture. It implements it.
How This Chapter Connects
This chapter extends the framework established in the preceding sections of the book:
- Chapters 1–12 demonstrated that Biblical Hebrew morphology is governed by a restricted control alphabet (10 letters, Z=152.16, p<0.0001). - \1 showed that the four divine names function as morphological state indicators with 87.8% predictive accuracy. - \1 (El Shaddai) traced the structural architecture of the Torah's narrative. - \1 established BovB/L1 equilibrium as the genomic signature of kashrut, with the sheep at unity and the nutrition cycle as a regulatory system. - \1 identified the Red Heifer as a genomic reference standard.
This chapter takes the next step: the same regulatory principle — balance, not complexity; management, not accumulation — distinguishes not only clean from unclean animals, but humans from all other primates. The control alphabet that governs Hebrew morphology, the divine names that mark regulatory states, the BovB/L1 ratio that classifies species, and the L1HS switch that enables human cognition are not four separate findings. They are four expressions of one architecture.
The following chapter returns to the Torah's narrative structure to examine how the parshiot (weekly readings) function as natural units within the statistical terrain — another layer of the same architecture, operating at the scale of text rather than genome.