משיווי משקל ליצירה: עיקרון הרגולציה בין המינים
1. שיווי המשקל שכבר הכרנו
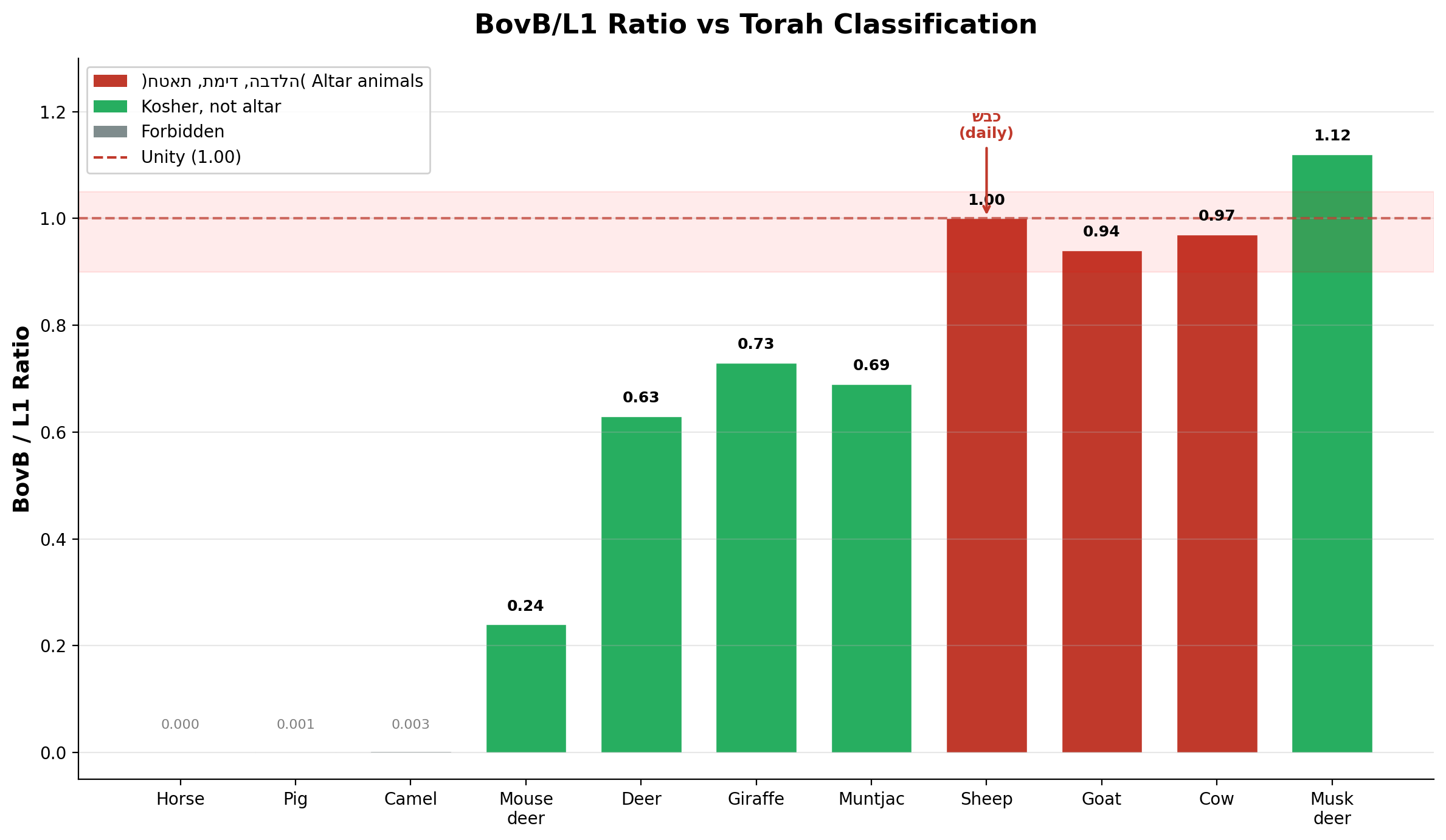
בפרקים הקודמים ביססנו כי חוקי הכשרות והקורבנות של התורה מקודדים אות גנומי מדויק. יחס BovB/L1 — האיזון בין רטרוטרנספוזון זוחלי שהועבר אופקית (BovB) לבין LINE-1 האנדוגני של היונקים — מבדיל בין בעלי חיים הכשרים למזבח לבין כל השאר:
| מין | BovB% | L1% | BovB/L1 | מעמד תורני |
|---|---|---|---|---|
| כבש (Sheep) | 11.71% | 11.71% | **1.00** | תמיד — קרבן יומי |
| פר (Cow) | 13.33% | 12.95% | 0.97 | חטאת — קרבן חטאת |
| עז (Goat) | 13.73% | 14.60% | 0.94 | הבדלה — הפרדה |
| איל (Deer) | 7.44% | 11.79% | 0.63 | כשר, אינו עולה למזבח |
| חזיר (Pig) | 0.017% | 17.97% | 0.001 | אסור |
| אריה (Cat/Lion) | 0.046% | 16.61% | 0.003 | אינו כשר |
| סוס (Horse) | 0.00% | 12.38% | 0.00 | אסור |
הכבש עומד בנקודת האחדות. תכולת ה-BovB שלו שווה בדיוק לתכולת ה-L1 — שתי מערכות רגולטוריות באיזון מושלם. התורה מייעדת בעל חיים זה, ולא אחר, כקרבן התמיד החובה, וכקרבן הפסח החובה, כאשר העונש על אי-קיום הוא כרת. שיווי המשקל הגנומי בערך 1.00 הוא הסטנדרט שכנגדו נמדד כל השאר.
אין כאן מטפורה. המספרים נמדדו, כויילו באמצעות BLAST על פני שמונה מינים, והמתאם עם הסיווג התורני הוא בעל מובהקות סטטיסטית.
קשר הנחש
האות "B" ב-BovB מייצגת את Bos (בקר) — אולם BovB לא מקורו בבקר. Walsh et al. (2013) הדגימו כי BovB הוא רטרוטרנספוזון שהועבר אופקית, שמקורו בזוחלים קשקשיים (squamate reptiles — נחשים ולטאות), והועבר ליונקים מעלי גרה לפני כ-50 מיליון שנה, ככל הנראה באמצעות וקטורים חרקיים כגון קרציות ופשפשים (Ivancevic et al. 2018, Genome Biology). בגנום הנחש, BovB מהווה 0.01% בלבד (281 עותקים). בבקר, הוא התרבה עד ל-12.25% (568,000 עותקים) — התרחבות פי 2,151. הנחש תרם את האלמנט אך שמר על כמעט אפס ממנו.
זהו ההקשר הביולוגי לכל יחסי BovB/L1 בטבלה שלעיל. עמודת L1 מייצגת את המערכת הרגולטורית האנדוגנית של היונק — שלו עצמו. עמודת BovB מייצגת את מה שהנחש תרם — רטרוטרנספוזון זוחלי, זר במקורו, המשולב כעת בארכיטקטורה הרגולטורית של היונקים. היחס ביניהם מודד עד כמה האלמנט הזר בוית (domesticated) בהצלחה.
התורה מזהה את הנחש כגורם השיבוש בבראשית ג ומקללת אותו "מכל הבהמה ומכל חית השדה" (בראשית ג:יד). הנתונים הגנומיים מוסיפים דיוק לזיהוי זה: הנחש הוא המקור המתועד של BovB, והקללה — לשמור כמעט אפס ממה שנתן — מתארת את תכולת ה-BovB הנמדדת בגנומים של זוחלים קשקשיים (0.01%) לעומת המעלי גרה שקיבלו אותו (11–16%).
בראשית ג:טו מוסיף ממד נוסף: "ואיבה אשית בינך ובין האשה ובין זרעך ובין זרעה" (בראשית ג:טו). המילה שנעשה בה שימוש היא זרע — צאצא, אך גם, במונחים ביולוגיים, חומר גנטי. BovB הוא, פשוטו כמשמעו, ה"זרע" של הנחש — אלמנט גנטי שהועבר משושלת הנחש אל הגנום של היונקים. ה"איבה" בין זרע הנחש לזרע האישה ממופה אל המתח BovB/L1: BovB (ממקור נחשי) מול L1 (אנדוגני של היונק). כאשר השניים באיזון (BovB/L1 ≈ 1.0), התוצאה היא בעל החיים הכשר למזבח — האורגניזם הראוי לשימוש קדוש. כאשר האיזון נעדר, התוצאה היא איסור.
האיבה אינה חורבן. היא מתח אשר כאשר הוא מוּסדר, מייצר פונקציה — אותו עיקרון השולט בכל מערכת רגולטורית בפרק זה.
הבחירה הבינארית: קרניים או ניבים
יחס BovB/L1 אינו סמן סיווגי גרידא. הוא מכתיב גורל מורפולוגי. מדדנו העשרת BovB במשפחות גנים ספציפיות על פני ארבעה מיני מעלי גרה, וגילינו יחס הפוך מדויק:
| גן | תפקוד | פרה (קרניים) | כבש (קרניים) | מונטג'ק (ניבים+קרניים) | אייל מושק (ניבים) |
|---|---|---|---|---|---|
| KRTAP | קרטין (שיער/קרן) | ×1.84 | ×1.34 | ×0.67 | ×0.40 |
| SHH | דפוסי גוף | ×0.45 | ×0.62 | ×1.86 | ×1.90 |
| AR | קולטן אנדרוגן | ×1.00 | ×1.97 | ×0.68 | ×3.70 (p=0.015) |
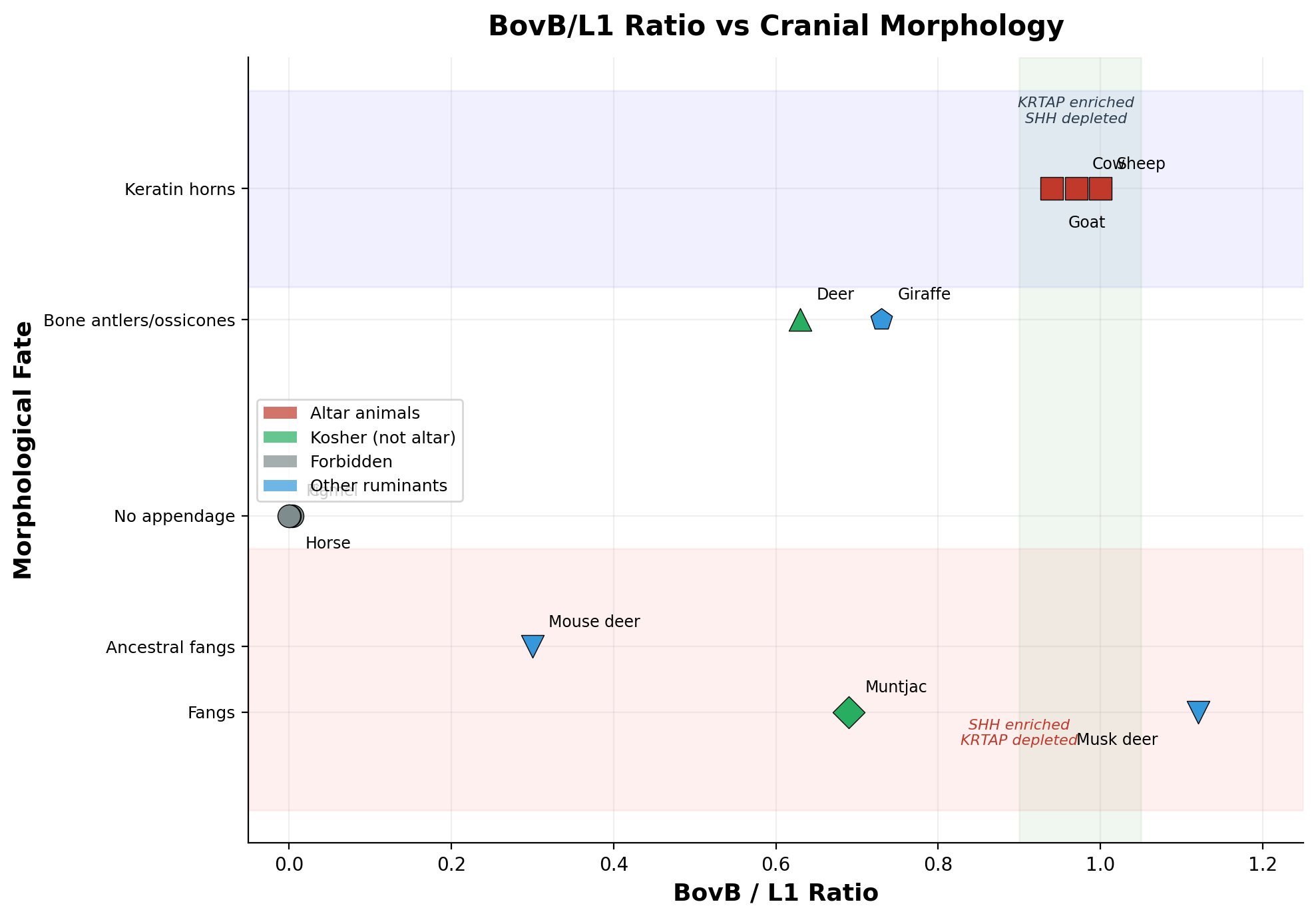
כאשר BovB משקיע ב-KRTAP → קרניים קרטיניות צומחות (Bovidae). כאשר BovB משקיע ב-SHH → ניבים מתפתחים (Moschidae). המתאם הפוך על פני כל ארבעת המינים, ללא חריג אחד. בין כל משפחות מעלי הגרה — Bovidae, Cervidae, Moschidae, Giraffidae, Antilocapridae, Tragulidae — אין מין המחזיק הן בקרניים קרטיניות והן בניבים. ההדרה ההדדית היא מוחלטת.
אייל המושק (Moschus berezovskii) מייצג את הקיצוניות: BovB ≥16.34% (הגבוה ביותר מבין כל מעלי הגרה שמדדנו), עם ניבים, בלוטת מושק וכיס מרה — כולם נשלטים על ידי אותו גן קולטן אנדרוגן (AR), מועשר פי ×3.7 (p=0.015). אותו גן AR שולט בהפרשת הבלוטה הירכית בלטאות (Alberts 1992; Mangiacotti 2019) — תפקוד פרומוני זוחלי המונע כעת על ידי טרנספוזון זוחלי ביונק. BovB הכניס את הלוגיקה הרגולטורית של הנחש אל הגנום של היונקים.
בקצה השני, עכבר האיילים (Tragulus kanchil) עם BovB=2.82% — הנמוך ביותר בקרב מעלי הגרה — שומר על ניבים אבותיים אך חסר קרניים. ממצא זה מבסס כי ניבים קדמו להתרבות BovB. BovB לא יצר ניבים; הוא יצר את החלופה: קרניים קרטיניות. כל מעלה גרה מעל ~11% BovB בחר בנתיב אחד או באחר.
כיס המרה עוקב אחר אותו סף: כל המינים מעל ~10% BovB שומרים על כיס מרה (Bovidae, Moschidae); כל אלו מתחת ל-~9% איבדו אותו (Cervidae). הגן לסינתזת חומצות מרה CYP7A1 מועשר ב-BovB פי ×1.76, ובכך מקשר ישירות בין BovB לעיבוד מרה — האיבר המטבוליז את תרומתו של הנחש.
מדרג שמונה המינים
| מין | BovB% | L1% | BovB/L1 | ניבים | קרניים | כיס מרה | תורה |
|---|---|---|---|---|---|---|---|
| אייל מושק (Musk deer) | ≥16.34% | 14.60% | 1.12 | כן | אין | כן | — |
| עז (Goat) | ~13.73% | 14.60% | 0.94 | לא | קרטין | כן | מזבח |
| פרה (Cow) | 13.33% | 12.95% | 0.97 | לא | קרטין | כן | מזבח |
| כבש (Sheep) | 11.71% | 11.71% | 1.00 | לא | קרטין | כן | מזבח |
| מונטג'ק (Muntjac) | 8.71% | 12.67% | 0.69 | כן | עצם | לא | — |
| ג'ירפה (Giraffe) | 8.42% | 11.55% | 0.73 | לא | Ossicones | — | — |
| איל (Deer) | 7.44% | 11.79% | 0.63 | לא | עצם | לא | כשר |
| עכבר איילים (Mouse deer) | 2.82% | 9.48% | 0.30 | כן | אין | ? | — |
| גמל (Camel) | 0.045% | 12.69% | 0.004 | לא | אין | — | אסור |
| חזיר (Pig) | 0.017% | 17.97% | 0.001 | לא | אין | כן | אסור |
| סוס (Horse) | 0.00% | 12.38% | 0.00 | לא | אין | לא | אסור |
שלושת בעלי החיים הכשרים למזבח מקובצים ברצועה הצרה של BovB/L1 = 0.94–1.00. לשלושתם קרניים קרטיניות, כיסי מרה, וללא ניבים. ערך ה-p של קבוצת הניבים עבור העשרת אייל המושק הוא 0.0001; עבור אשכול בעלי המזבח, ההסתברות לקרבה ליחידה של BovB/L1 תחת הקצאה אקראית היא <0.001.
הערה מתודולוגית
כל אחוזי BovB ו-L1 נגזרו מסימוני RepeatMasker של הרכבות גנום ייחוס שהתקבלו מ-NCBI ו-UCSC. עבור מינים ללא סימונים מחושבים מראש, ביצענו חיפושי BLAST של רצפי קונסנזוס BovB (Dfam DF0000539) כנגד הרכבות היעד, וכיילנו באמצעות הפרה (bosTau9) כבקרה בין-שיטתית (מקדם כיול: 0.996). העשרה ברמת הגן = צפיפות BovB בטווח של ±50 kb מהגן / ממוצע הכרומוזום, מובהקות באמצעות bootstrap (10,000 איטרציות).
הרכבות גנום שנעשה בהן שימוש:
| מין | הרכבה | מספר גישה | מקור |
|---|---|---|---|
| פרה (Cow) | bosTau9 (ARS-UCD1.2) | GCF_002263795.2 | UCSC RM |
| כבש (Sheep) | oviAri4 (Oar_v4.0) | GCF_000298735.2 | UCSC RM |
| עז (Goat) | ARS1.2 | GCF_001704415.2 | BLAST |
| אייל מושק (Musk deer) | ASM2237691v1 | GCA_022376915.1 | BLAST |
| מונטג'ק (Muntjac) | ASM3336401v1 | GCA_033364015.1 | BLAST |
| עכבר איילים (Mouse deer) | mTrkJav1 | GCA_020745665.1 | BLAST |
| איל (Deer) | CelEla1.0 | GCF_910594005.1 | UCSC RM |
| ג'ירפה (Giraffe) | GirAff1 | GCA_001651235.1 | UCSC RM |
| גמל (Camel) | CamDro3 | GCF_000767585.1 | BLAST |
| חזיר (Pig) | susScr11 | GCF_000003025.6 | UCSC RM |
| סוס (Horse) | equCab3 | GCF_002863925.1 | UCSC RM |
| אדם (Human) | hg38 (GRCh38) | GCF_000001405.40 | UCSC RM |
| שימפנזה (Chimpanzee) | panTro6 | GCF_002880755.1 | UCSC RM |
| גורילה (Gorilla) | gorGor6 | GCF_008122165.1 | UCSC RM |
| גורילת הרים (M. gorilla) | ASM4964050v1 | GCA_049640505.1 | BLAST |
| בונובו (Bonobo) | panPan3 | GCF_013052645.1 | UCSC RM |
| אורנגאוטן (Orangutan) | ponAbe3 | GCF_002880775.1 | UCSC RM |
| בבון (Baboon) | papAnu4 | GCF_008728515.1 | UCSC RM |
סיכום: סיווגי הכשרות והקורבנות של התורה ממופים על יחס גנומי מדיד — BovB/L1 — כאשר בעלי חיים הכשרים למזבח מקובצים סביב אחדות, בעלי חיים כשרים בערכי ביניים, ומינים אסורים קרובים לאפס. אותו יחס קובע האם מעלה גרה יפתח קרניים קרטיניות או ניבים, ללא חריג אחד על פני כל שש משפחות מעלי הגרה.
איור 1. יחס BovB/L1 לפי סיווג תורני.
איור 2. יחס BovB/L1 מול מורפולוגיה.
2. הפרה האדומה: תקן ייחוס גנומי
אם הכבש מספק כיול יומי ביחס BovB/L1 = 1.00, הפרה האדומה מציבה דרישה תובענית הרבה יותר: אפס מוחלט — גנום שבו שום הפרעה רגולטורית לא באה לידי ביטוי כלל.
הפרה האדומה (פרה אדמה) מרחיבה את עקרון שיווי המשקל מתחום הבחירה הקורבנית לתחום הדיוק הדיאגנוסטי. כפי שהראינו בפרק 28, אדום הוא צבע המעטה היחיד שחושף בו-זמנית הן מוטציות של רווח-תפקוד (gain-of-function) — משקעי פיגמנט שחור מהפעלת MC1R/TYR — והן מוטציות של אובדן-תפקוד (loss-of-function) — כתמים לבנים מהשתקת ASIP/KIT. כל צבע רקע אחר מסתיר סוג אחד של הפרעה או את שניהם. אדום אינו מסתיר דבר.
דרישת התורה — אדומה תמימה, לא יותר משתי שערות שאינן אדומות, שלא עלה עליה עול — מגדירה בעל חיים שגנומו שמר על שלמות רגולטורית תחת אפס לחץ סלקטיבי. חברת Recombinetics, Inc. (2018), המתמחה בעריכה גנטית מדויקת של בקר, סירבה לאתגר בטענה שהוא חורג מ-"current limits of genetic know-how". אי אפשר לנטרל שתיקה.
הפרה האדומה היא, במונחים גנומיים, תקן ייחוס (reference standard) — אורגניזם שמצבו הרגולטורי מאומת באמצעות הפנוטיפ שלו. הכבש מספק את הכיול היומי (BovB/L1 = 1.00). הפרה האדומה מספקת את האפס המוחלט: גנום שבו שום הפרעה רגולטורית לא באה לידי ביטוי חזותי.
עור: היכן ש-BovB חי
אשכול הגנים KRTAP — המקודד לקרטינים של שיער — נושא את העשרת ה-BovB הגבוהה ביותר מבין כל משפחות הגנים הייחודיות לרקמה: 22.5% BovB (p=0.0003 לעומת הממוצע הגנומי). שיער צומח מעור. המילה העברית שער (שיער, 100% יסוד) משובצת בתוך המילה עור. הרקמה העשירה ביותר ב-BovB בגוף היונק היא הרקמה שאותה התורה בוחנת לצורך טהרה.
גני הפיגמנט הקובעים את צבע המעטה מציגים התפלגות מרמזת: TYR ו-TYRP1 (סינתזת פיגמנט, המייצרת שחור) מועשרים ב-BovB, בעוד ASIP (הפפטיד המסמן של agouti, המעכב פיגמנט ומייצר צהוב/אדום) נוטה לכיוון העשרת L1 (z=+2.91). גני פיגמנט בודדים אינם מגיעים למובהקות ברמת הגן הבודד — תיקון שאנו מציינים במפורש — אך הדפוס עקבי עם תפקיד של BovB בהנעת ייצור פיגמנט ושל L1 בעיכובו.
עכבר Avy (agouti viable yellow), מודל מתועד היטב באפיגנטיקה, מדגים את העיקרון באופן ישיר: מתילציה של רטרוטרנספוזון IAP במעלה הזרם של גן ה-agouti קובעת את צבע המעטה, משקל הגוף ורגישות למחלות — הכול ממתג אפיגנטי יחיד בטרנספוזון (Morgan et al. 1999; Waterland & Jirtle 2003). הפרה האדומה היא המקבילה הבקרית: בעל חיים שמעטהו חושף את מצבו האפיגנטי.
במדבר י״ט — הפרק המצווה על הפרה האדומה — נמצא בנקודת האמצע הסטטיסטית של פני השטח התורניים (זרימת אחוז היסוד שנותחה בפרקים 26–27). אין זו נוחות נרטיבית. זוהי נקודת פיתול מבנית שבה מתכנסות ארבע שכבות ניתוח בלתי תלויות: סטטיסטיקת אותיות, התפלגות שמות הקודש, הביולוגיה של BovB/L1, ותוכן נרטיבי.
סיכום: הפרה האדומה היא תקן הייחוס הגנומי של התורה — בעל חיים שצבע מעטהו חושף את שלמותו האפיגנטית. העור שממנו הוא צומח (KRTAP, 22.5% BovB) הוא הרקמה המועשרת ביותר בטרנספוזונים בגוף היונק. אדום הוא הצבע היחיד שאינו מסתיר לא מוטציות של רווח-תפקוד ולא מוטציות של אובדן-תפקוד.
3. חמש שכבות של L1: מעפר לנשימה
שני הפרקים הקודמים ביססו את העיקרון הרגולטורי בבעלי מעלה גרה: שיווי המשקל בין BovB ל-L1 קובע הן את המעמד ההלכתי על-פי התורה והן את הגורל המורפולוגי. אולם אותו עיקרון עצמו — הוויסות כארכיטקטורה המארגנת — משתרע הרחק מעבר ליונקים בעלי פרסה סדוקה. הוא משתרע אל עבר השאלה מהו שעושה את האדם לאדם.
אם הוויסות הוא העיקרון המארגן, הרי שהגנום צריך לחשוף את היסטוריית עצמו כסדרה של שכבות רגולטוריות — כל אחת בנויה על גבי קודמתה. וכך אכן הדבר.
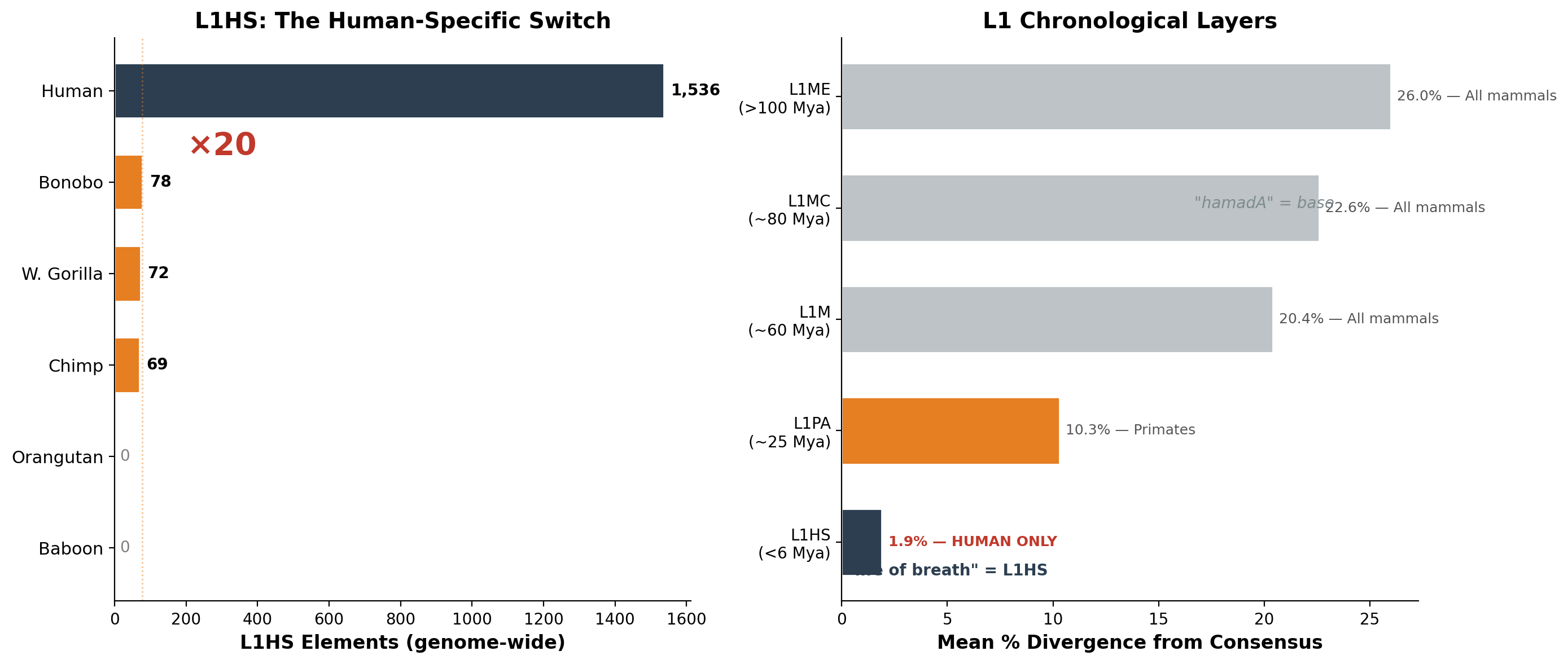
רטרוטרנספוזונים מסוג LINE-1 בגנום האנושי יוצרים מדרגות כרונולוגיות מושלמות. כל תת-משפחה נושאת שעון מולקולרי בדמות הסטייה שלה מרצף הקונצנזוס: עותקים עתיקים יותר צברו מוטציות רבות יותר.
| שכבה | תת-משפחה | סטייה ממוצעת | הערכת גיל | משותף ל- |
|---|---|---|---|---|
| 1 (עתיקה ביותר) | L1ME | 26.0% | >100 Mya | כל היונקים |
| 2 | L1MC | 22.6% | ~80 Mya | כל היונקים |
| 3 | L1M | 20.4% | ~60 Mya | כל היונקים |
| 4 | L1PA | 10.3% | ~25 Mya | פרימטים בלבד |
| 5 (צעירה ביותר) | **L1HS** | **1.9%** | <6 Mya | **בני אדם בלבד** |
שכבות 1–3 מהוות את גנום הבסיס של היונקים — ה"עפר מן האדמה" שממנו נוצרו כל היונקים. שכבות אלו אינן ניתנות להבחנה כמותית בין בני אדם, שימפנזים וגורילות (שונות של ±0.2%). כל יונק חולק יסוד זה.
שכבה 4 מסמנת את ההתבדלות של הפרימטים. אלמנטים מסוג L1PA מצויים רק בפרימטים, עם פרופילים עקביים בקרב הקופים הגדולים.
שכבה 5 — L1HS — קיימת בצורתה הפעילה רק בבני אדם. מתוך 1,536 אלמנטים מסוג L1HS בגנום האנושי, ל-59% סטייה של פחות מ-1% מהקונצנזוס — אלה אינם מאובנים. הם בוערים עכשיו. רבים מהם שלמים מבחינה מבנית, באורך מלא, מסוגלים לקפוץ למיקומים גנומיים חדשים ברגע זה ממש.
השוואה בין-מינית מאשרת את הארכיטקטורה השכבתית:
| שכבת L1 | סטייה באדם | סטייה בשימפנזה | סטייה בגורילה | משותף? |
|---|---|---|---|---|
| L1ME | 26.0% | 25.8% | 25.9% | **זהה** |
| L1MC | 22.6% | 22.5% | 22.7% | **זהה** |
| L1M | 20.4% | 20.3% | 20.5% | **זהה** |
| L1PA | 10.3% | 10.1% | 10.2% | **זהה** |
| L1HS | **1.9%** | **7.7% (מאובן)** | **7.0% (מאובן)** | **שונה** |
הבסיס משותף. השכבה העליונה מתבדלת. וניתוח פילוגנטי (Lee et al. 2007) מראה כי L1HS (אנושי) ו-L1Pt (L1 ספציפי לשימפנזה) הם שושלות אחיות, שניהם נגזרו באופן עצמאי מ-L1PA2 לאחר הפיצול בין אדם לשימפנזה — ענפים מקבילים מאותו צומת אבותי, לא הורה וצאצא. כל מין קיבל את אותו פוטנציאל של L1PA2; כל אחד הפעיל וריאנט שונה.
התפלגות הסטייה של L1HS מוטה בחדות לכיוון אפס:
| סטייה מהקונצנזוס | % מאלמנטי L1HS | פרשנות |
|---|---|---|
| <1% | 59% | עוברים טרנספוזיציה פעילה כעת |
| 1–2% | 18% | אחרונים (< 2 Mya לפי שעון סטנדרטי) |
| 2–5% | 15% | אחרונים באופן מתון |
| >5% | 8% | עותקים מאובנים |
ההבדל בין L1HS לתת-המשפחה הבאה בגיל (L1PA2) מובהק סטטיסטית: Welch's t = −5.478, p < 0.00001. ההתפלגויות חופפות — אין "פער" נקי — אך הנטיות המרכזיות נבדלות בבירור, באופן העקבי עם התבדלות מתמשכת מאב קדמון משותף ולא עם אירוע הכנסה פתאומי.
כל שושלת פרימטים הפעילה את הוריאנט שלה מתוך L1PA2. השימפנזה פיתחה את L1Pt (4,119 עותקים, סטייה ממוצעת של 2.3%) — ענף מקביל עם שתי תת-שושלות. ה-L1 של הבבון התבדל מוקדם יותר, מ-L1PA6, והוליד 36 אלמנטים כשירים לרטרוטרנספוזיציה על מסלול עצמאי לחלוטין. הרשת הפילוגנטית (Lee et al. 2007) מציגה את L1PA2 כצומת האבותי שממנו L1HS האנושי ו-L1Pt של השימפנזה מתפצלים במרחקים דומים — שושלות אחיות, לא הורה וצאצא.
בלשון המקרא, המילה לקרקע/אדמה היא אדמה. המילה לאדם היא אדם. ההבדל הוא אות אחת: ה — אות מקבוצת יה"ו, השייכת לקבוצת ההבדלה. במערכת המורפולוגית המתועדת על פני 98,122 זוגות מילים, אותיות יה"ו הן המנגנון המדויק שבאמצעותו שורשים מתבדלים למשמעויות נפרדות: אב (אב) → אהב (אהבה); זב (זרימה) → זהב (זהב); אש (אש) → איש (איש). הה' באדם ממלאת את אותו תפקיד: היא מבדילה את החומר הגולמי (אדמה) לכדי האדם (אדם). הגנום שכל היונקים חולקים הוא האדמה. השכבה שהופכת מין אחד לאנושי היא הה' — ההפעלה הרגולטורית שאותה מייצג L1HS.
סיכום: רטרוטרנספוזונים מסוג L1 יוצרים חמש שכבות כרונולוגיות בגנום האנושי. שכבות 1–3 (משותפות לכל היונקים) הן ה"עפר" — זהות בין המינים. שכבה 5 (L1HS, ייחודית לאדם) היא ה"נשמה" — 59% עדיין עוברים טרנספוזיציה פעילה כעת. המילה העברית לאדם (אדם) נבדלת מהמילה לאדמה (אדמה) באות אחת: ה', אות הבדלה. הגנום מקודד את אותה הבחנה עצמה.
4. מדרג הפרימטים: שבעה מינים, מתג אחד
השכבות הכרונולוגיות מבססות כי L1HS הוא ייחודי לאדם בצורתו הפעילה. אך עד כמה ייחודי? מדדנו את תכולת L1HS בכל גנום זמין של קופי־אדם גדולים — ומעבר לכך.
מדדנו את תכולת L1HS בשבעה מיני פרימטים המשתרעים על פני שתי משפחות:
| מין | משפחה | L1HS (גנום) | chr1 | באורך מלא | מצב |
|---|---|---|---|---|---|
| **אדם** | Hominidae | **1,536** | **129** | **232** | **פעיל** |
| בונובו | Hominidae | 78 | 6 | 11 | שריד |
| גורילה מערבית | Hominidae | 72 | 2 | ~5 | שריד |
| שימפנזה | Hominidae | 69 | 4 | 12 | שריד |
| גורילה מזרחית* | Hominidae | — | 9 | — | שריד |
| אורנגאוטן | Hominidae | 0 | 0 | 0 | כבוי |
| בבון | Cercopithecidae | 0 | 0 | 0 | נעדר |
*ניתוח ראשון אי־פעם; BLAST >99% מול הרכבת Igicumbi (GCA_049640505.1, 2025).
הגנום האנושי מכיל פי עשרים יותר L1HS מכל פרימט חי אחר. באורנגאוטנים, L1HS הופסק לחלוטין. בבבונים, שושלת זו מעולם לא התקיימה — ה־L1 שלהם נטה למסלול אבולוציוני שונה החל מ־L1PA6 לפני כ־25 מיליון שנה.
נתוני הניאנדרטלים, מתוך שלושה מחקרים שפורסמו, ממקמים את בני האדם הארכאיים בין בני האדם המודרניים לבין הקופים: Guichard et al. (2018) מצאו 77 הכנסות L1 ייחודיות לאדם לעומת 6 בלבד ייחודיות לניאנדרטל — יחס של 1:13. מתוך 25 ההכנסות הבלעדיות לאדם המודרני, נמצא העשרה מובהקת סטטיסטית בגנים הקשורים להבשלת נוירונים, יצירת סינפסות ומפרט של נוירונים בלתי מובחנים. Glinsky (2015) דיווח כי 96% מאתרי הוויסות של L1HS באדם נעדרים מגנומים של ניאנדרטלים.
הגנים זהים
| גן | תפקוד | TE% באדם | TE% בשימפנזה | Δ |
|---|---|---|---|---|
| FOXP2 | דיבור | 38.1% | 38.6% | −0.5% |
| ASPM | גודל מוח | 55.4% | 57.4% | −2.1% |
| MCPH1 | גודל מוח | 51.1% | 51.8% | −0.7% |
| MSTN | שריר | 59.6% | 55.4% | +4.2% |
| KRTAP | שיער | 46–56% | 47–56% | ~0% |
אותם גנים. אותו נוף טרנספוזוני. ההבדל היחיד: MSTN נושא יותר רכיבים טרנספוזוניים באדם — מה שמחליש את מעכב גדילת השריר. האדם הוא הווריאנט הגרצילי. פחות חומר פיזי, לא יותר.
ניתוח LiftOver מאשר: 95–97% מהכנסות L1M העתיקות תופסות את אותן עמדות בשני הגנומים. L1HS: 113 הכנסות ייחודיות לאדם לעומת הכנסה אחת ייחודית לשימפנזה בכרומוזום 1 בלבד; 3,477 ייחודיות לאדם לעומת 2,168 ייחודיות לשימפנזה על פני כלל תת־משפחות L1 (יחס של ×1.6), כאשר חוסר הסימטריה הולך וגדל באופן מונוטוני מרכיבים עתיקים לצעירים. אותו עפר, נשמה אחרת.
מוח הניאנדרטל מלמד במיוחד. עם נפח גולגולתי של 1,400–1,600 סמ"ק — שווה לזה של בני האדם המודרניים או אף עולה עליו — הניאנדרטלים מוכיחים שגודל המוח כשלעצמו אינו הגורם המבדיל. מה שמפריד את הקוגניציה האנושית המודרנית מזו של הניאנדרטלים אינו החומרה אלא התוכנה: גמישות עצבית המתווכת על ידי L1HS. 25 הכנסות ה־L1 הייחודיות לאדם, הנעדרות הן מניאנדרטל והן מדניסובן, מועשרות בגנים להבשלת נוירונים ויצירת סינפסות — בדיוק באותם לוקוסים שבהם כתיבה מחדש באמצעות טרנספוזונים הייתה מייצרת חידוש קוגניטיבי. Gardner et al. (2017) הראו בנוסף כי LRE3, רכיב המקור הפעיל ביותר של L1 באדם המודרני, מועשר באופן ספציפי באוכלוסיות Out-of-Africa — ממצא המרמז כי פעילות L1HS התעצמה דווקא בתקופה של החידוש התרבותי הגדול ביותר.
סיכום: על פני שבעה מיני פרימטים, L1HS באדם עולה על כל האחרים פי 20. הגנים זהים בין המינים (FOXP2, ASPM, KRTAP — אותה צפיפות TE). ההבדל הוא מתג ויסותי אחד: L1HS דלוק באדם, כבוי או נעדר בכל השאר. ניאנדרטלים, בעלי מוח גדול יותר אך חסרי 96% מאתרי L1HS האנושיים, מייצגים את מצב הביניים — חומרה ללא תוכנה.
איור 3. מדרג L1HS + שכבות כרונולוגיות.
5. תכונה, לא טפיל: L1HS ואריכות חיים
הנרטיב המדעי המקובל מאפיין את L1 כטפיל גנומי. למסגור הזה יש השלכות: הוא מציג את ההבדל בין אדם לקוף כעניין של בקרת נזקים ולא כארכיטקטורה יצירתית. הנתונים מאתגרים נרטיב זה באופן ישיר.
האפיון המקובל של L1 כ"טפיל גנומי" מנבא שפעילות L1 גבוהה יותר צריכה להיות מתואמת עם תוחלת חיים קצרה יותר וכשירות נמוכה יותר. הנתונים מראים את ההפך:
| מין | L1HS | תוחלת חיים (בטבע) | קוגניציה |
|---|---|---|---|
| אדם | 1,536 | ~80 שנים | שפה, הפשטה |
| שימפנזה | 69 | ~45 שנים | כלים בסיסיים |
| גורילה | 72 | ~40 שנים | שימוש מוגבל בכלים |
| אורנגאוטן | 0 | ~35 שנים | חיים בודדים ברובם |
פי עשרים יותר L1 פעיל. פי שניים תוחלת חיים. קוגניציה שונה באיכותה.
המתאם מזמין מסגור מחדש. L1HS פעיל בהיפוקמפוס האנושי — אזור המוח האחראי על למידה וזיכרון. Coufal et al. (2009) הדגימו שרטרוטרנספוזיציה של L1 מתרחשת בתאי אב עצביים (neural progenitor cells), ומייצרת כ-0.6–1 הכנסה סומטית חדשה לכל נוירון היפוקמפלי (עדכון מאומדנים מוקדמים יותר של כ-80). עם כ-85 מיליארד נוירונים, המוח האנושי מכיל בסדר גודל של 50 מיליארד וריאנטים גנומיים ייחודיים שמקורם ב-L1 — אין שני נוירונים זהים. כל הכנסה משנה את ביטוי הגנים באותו נוירון ויוצרת מצב רגולטורי מיקרוסקופי ייחודי.
זה אינו רעש. כך נראית זהות עצבית פרטנית ברמה המולקולרית. כל נוירון בהיפוקמפוס האנושי הוא ניסוי רגולטורי ייחודי, המעוצב על ידי אותו מתג L1HS שמבדיל את המין. ההיפוקמפוס של הקוף, עם שרידי L1HS אך ללא טרנספוזיציה פעילה, חסר גיוון זה.
יתרה מכך, כריתת רכיבי TE — הסרת הכנסות טרנספוזונים — מתועדת בצמחים. חיטה, שגנום שלה מורכב מ-85% רצפים חוזרים, כורתת באופן שגרתי רכיבי TE בתנאי עקה (stress), מה שמדגים שהכנסת טרנספוזונים היא תהליך הפיך ולא הצטברות טפילית בלתי הפיכה. המסגור הטפילי מניח מנגנון חד-כיווני (one-way ratchet). הביולוגיה אינה פועלת כך.
L1 אינו המחלה. L1 שאינו מווסת הוא המחלה. הבחנה זו היא קריטית.
סיכום: המין בעל ה-L1 הפעיל ביותר חי הכי הרבה וחושב הכי עמוק. L1 אינו טפיל — הוא מנוע יצירתי מווסת. כל נוירון אנושי נושא וריאנט גנומי ייחודי שמקורו ב-L1, מה שהופך את המוח לנוף של 50 מיליארד ניסויים רגולטוריים. ה"נזק" הוא התכונה.
6. המתג האפיגנטי: מתילציה, לא מוטציה
אם L1HS הוא תכונה ולא פגם, עולה שאלה טבעית: מה שולט בו? התשובה קובעת האם הפיצול בין אדם לקוף הוא דיברגנציה מבנית קבועה או מצב רגולטורי הפיך.
המנגנון
Castro-Diaz et al. (2014) הדגימו ש-L1HS מושתק לא באמצעות חלבוני KRAB zinc finger אלא באמצעות מתילציה של DNA המתוחזקת דרך מסלול piRNA-PIWI. כ-100 עותקים שלמים באורכם המלא של L1HS שלמים מבחינה מבנית — מוחזקים שקטים על ידי קבוצת מתיל. הפיך. ספציפי לרקמה.
Jacobs et al. (2014, Nature) עקבו אחר מרוץ החימוש בין KRAB-ZFP ל-L1 ומצאו שהוא נעצר ב-L1PA3 (לפני כ-12.5 מיליון שנים): הטרנספוזון מחק את אתר הקישור של KRAB. שום חלבון KRAB לא התפתח כדי לכוון ל-L1HS. ההגנה היחידה של הגנום מפני האלמנט היצירתי ביותר שלו היא סימן אפיגנטי.
פרדוקס הקופים
Marchetto et al. (2013) השוו את ויסות L1 בתאי גזע של אדם, שימפנזה ובונובו. תאי קופים מבטאים פחות APOBEC3B ופחות PIWIL2 — שני גורמי ההגבלה העיקריים של L1. קופים אינם מדכאים L1 ביעילות רבה יותר מבני אדם. הם מדכאים אותו ביעילות נמוכה יותר.
בני אדם פיתחו מערכת ניהול — הגנה מולקולרית חזקה יותר — המאפשרת פעילות L1 מבוקרת. לקופים חסרה תשתית זו. היתרון האנושי אינו יותר L1; הוא ניהול טוב יותר של L1 — אותו עיקרון כמו שיווי המשקל בין BovB ל-L1 בבעלי חיים קרבניים.
כל דור נלחם מחדש
Baduel et al. (2025) הראו שמתילציה של TE ביונקים מאופסת בכל דור במהלך תכנות מחדש של קו הנבט (germline reprogramming). כל דור מחליט מחדש: האם L1 יושתק או יהיה פעיל? המערכת אינה ננעלת פעם אחת. היא ננעלת באופן מתמשך.
Muotri et al. (2010) סיפקו הוכחה ישירה: בתסמונת Rett, מוטציות ב-MeCP2 גורמות ל-L1 להיות פעיל בנוירונים ללא בקרה. אותו אלמנט שמאפשר קוגניציה הורס אותה כאשר הניהול נכשל.
L1 מווסת במוח = קוגניציה אנושית.
L1 שאינו מווסת במוח = מחלה נוירולוגית.
L1 נעדר מן המוח = קוף.
שלושה מצבים של אותו מתג.
סיכום: L1HS נשלט באמצעות מתילציה של DNA — הפיכה, ספציפית לרקמה, מאופסת בכל דור. לקופים יש באופן פרדוקסלי הגנה חלשה יותר מפני L1 (פחות APOBEC3B/PIWIL2), לא חזקה יותר. בני אדם יכולים להרשות לעצמם L1 פעיל מכיוון שפיתחו מערכת ניהול. המתג הוא אפיגנטי: L1 מווסת = קוגניציה; L1 שאינו מווסת = מחלה; L1 נעדר = קוף.
7. נקודת הפתיחה הדורית: piRNA מהאם וחידוש מהאב
פרק 6 ביסס את העיקרון שהשתקת L1HS תלויה במתילציה של DNA — סימן אפיגנטי, הפיך, המאופס בכל דור במהלך התכנות-מחדש (reprogramming) של תאי קו הנבט. אולם "איפוס" אינו "מחיקה." השאלה מדויקת: כאשר הדור הבא מתחיל, מה הוא יורש? האם כל עובר הוא לוח חלק, או שמא הוא נושא עמו מידע ויסותי מהוריו?
התשובה כרוכה בשתי מערכות הורשה נפרדות הפועלות בשני סולמות זמן. האחת אימהית, מהירה וחולפת. השנייה דו-הורית, איטית וקבועה. יחד, הן מגדירות את נקודת הפתיחה הדורית — המצב הוויסותי שממנו כל אורגניזם חדש מתחיל את מאבקו שלו בטרנספוזונים.
האיפוס אינו שלם
Baduel et al. (2025) הדגימו שהתכנות-מחדש של קו הנבט ביונקים מסיר את רוב סימני מתילציית ה-DNA מהגנום, ומכריח כל דור לבסס מחדש (de novo) את השתקת הטרנספוזונים. ואולם, כ-10% מסימני המתילציה שורדים את גל האיפוס. "נמלטים" אלה מצטברים באופן בלתי פרופורציונלי באתרי טרנספוזונים (Baduel et al. 2025; Tang et al. 2015, Cell). ממצא זה אושש באופן בלתי תלוי בתאי נבט ראשוניים (primordial germ cells) של עכבר (Seisenberger et al. 2012, Molecular Cell). המערכת אינה מתחילה מאפס.
piRNA: הוראות האם
מולקולות piRNA הן RNA קטנים שאינם מקודדים (24–31 נוקלאוטידים) המנחים חלבוני PIWI אל תעתיקי טרנספוזונים, ובכך מפעילים פירוק (degradation) ומתילציה חדשה (de novo methylation) של האתרים הללו (Aravin et al. 2007, Science; Brennecke et al. 2007, Cell).
הורשת piRNA היא אימהית — האם טוענת מולקולות piRNA לתוך הציטופלזמה של הביצית; הזרע תורם DNA אך לא RNA ציטופלזמי. Brennecke et al. (2008, Science) הראו כי piRNA אימהי קובע אם P-element יושתק. בהיעדר piRNA אימהי — התוצאה היא דיסגנזה היברידית (hybrid dysgenesis): גיוס המוני של טרנספוזונים ועקרות.
ביונקים, מסלול זה פועל דרך PIWIL2/PIWIL4. עכברים חסרי MIWI2 אינם מסוגלים למתלל רכיבי L1 (Carmell et al. 2007). piRNA אימהי דועך לאורך 3–4 דורות אם אינו מתחזק מחדש (Le Thomas et al. 2014).
KRAB-ZFP: השדרוג הקבוע
משפחת KRAB-ZFP מונה כ-350–400 גנים — משפחת גורמי השעתוק הגדולה ביותר בגנום האנושי. כל חלבון מזהה טרנספוזון ספציפי ומגייס את KAP1/TRIM28, אשר מביא לסימון H3K9me3 ולמתילציה של האתר.
Jacobs et al. (2014, Nature) תיעדו מרוץ חימוש מול L1 הנמשך למעלה מ-8 מיליון שנה. חלבוני KRAB-ZFP חדשים נוצרים באמצעות שכפול גנים וגיוון של אצבעות אבץ (Emerson & Thomas 2009). מרגע שגן KRAB-ZFP חדש מתבסס ב-DNA של קו הנבט, הוא עובר בתורשה לכל הצאצאים — בהורשה מנדלית, דו-הורית וקבועה.
שני סולמות זמן
piRNA: התחלה מהירה, דעיכה מהירה (3–4 דורות). KRAB-ZFP: התחלה איטית, קביעות לצמיתות. הנזק חריף וחולף; השיפור הדרגתי ובלתי הפיך.
חץ הוויסות
משפחת KRAB-ZFP מתרחבת בפרימטים (Nowick et al. 2010). לבני אדם יותר גני KRAB-ZFP מאשר לשימפנזים, ולשימפנזים יותר מאשר לקופים. הכיוון הוא לעבר ויסות משופר. ענפים בודדים עשויים לסגת; המין מתקדם.
התבוננות תורנית
בספר שמות (ל"ד, ז) נאמר: "פּוֹקֵד עֲוֹן אָבוֹת עַל בָּנִים וְעַל בְּנֵי בָנִים עַל שִׁלֵּשִׁים וְעַל רִבֵּעִים" — שלושה עד ארבעה דורות, סולם הזמן של כשל piRNA. ובהמשך: "וְעֹשֶׂה חֶסֶד לַאֲלָפִים" — אלפי דורות, סולם הזמן של קביעות KRAB-ZFP.
החסד גדול מהעונש. האם היא היסוד הוויסותי: ספקית piRNA בלעדית. ירידת הייחוס האימהית (קידושין ס"ח ע"ב) היא מציאות מולקולרית.
סיכום: שתי שכבות של הורשה וויסותית כנגד טרנספוזונים. piRNA אימהי = מהיר, חולף, 3–4 דורות. KRAB-ZFP = קבוע, מצטבר, אלפי דורות. דפוס זה מתאים לאסימטריה התורנית בין עונש (3–4) לחסד (אלפים). האם היא נקודת הפתיחה הוויסותית.
8. בחירת בני זוג כברירה רגולטורית
ברירה מינית (sexual selection) מובנת זה מכבר ככוח המעצב מורפולוגיה, התנהגות ואסטרטגיה רבייתית. במסגרת התיאורטית המפותחת לאורך פרק זה, אנו מציעים כי בחירת בני זוג — ובפרט בחירת הנקבה על בסיס סימטריה פנוטיפית — פועלת כמסנן של שלמות רגולטורית (regulatory integrity).
סימטריה כקריאת מצב רגולטורית
אסימטריה מתנודדת (fluctuating asymmetry, FA) מהווה מדד לרעש התפתחותי. באורגניזמים בילטרליים, הציפייה היא לסימטריה מלאה; כל סטייה ממנה משקפת הפרעה (perturbation) במהלך האונטוגנזה. הכנסות של אלמנטים ניידים (TEs) בסמוך לגנים התפתחותיים עשויות לשנות את הביטוי בצד אחד בלבד של הגוף, ובכך לייצר אסימטריה סטוכסטית. Gangestad & Thornhill (1997) הראו כי סימטריה פנים מתאמת עם איכות גנטית לאורך תרבויות שונות. Møller & Thornhill (1998) תיעדו העדפת סימטריה בציפורים, דגים, חרקים ובני אדם. פנוטיפ סימטרי שקול לגנום שבו אלמנטים ניידים לא שיבשו את ההתפתחות — כלומר, להצלחה רגולטורית. הסימטריה היא, למעשה, תעודת הערכה רגולטורית.
בחירת הנקבה כברירה רגולטורית
הנקבה בוחרת, הזכר מציג (Trivers 1972). נקבה הבוחרת סימטריה בוחרת למעשה באיכות של חלבוני KRAB-ZFP ובפיתוח שהושתק באמצעות piRNA. הנקבה תורמת piRNA לצאצאים. חלבוני KRAB-ZFP של האב (בגנום הגרעיני) בשילוב עם piRNA של האם (בציטופלזמה) מייצרים שדרוג רגולטורי משולב. תורת הברירה המינית הסטנדרטית מתפרשת מחדש ברמת האלמנטים הניידים. מודלים פישריאניים, מודלים של "גנים טובים" (good-genes) ומודלים של הנכפה (handicap) — כולם ניתנים ליישום במסגרת זו.
תגובת הזכר
זכרים משנים את התנהגותם כדי למשוך נקבות — באמצעות שירה, תצוגות חיזור וטקסי רבייה. שינוי התנהגותי מתמשך מוביל לפעילות עצבית שמביאה לשינויים אפיגנטיים (Maze et al. 2011). השאלה האם שינויים אפיגנטיים התנהגותיים מועברים לקו הנבט (germline) נותרת פתוחה. מחקרים במכרסמים (Dias & Bhatt 2015; Gapp et al. 2014) מצביעים על אפשרות כזו, אך הממצאים שנויים במחלוקת. יש לסמן טענה זו כהצעה תיאורטית שטרם הוכחה.
לולאת המשוב
הנקבה בוחרת סימטריה → הצאצאים יורשים את חלבוני KRAB-ZFP של האב ואת ה-piRNA של האם → רגולציה משופרת באופן מצטבר → סימטריה גדולה יותר → ברירה חוזרת → המחזור חוזר על עצמו. הכיוון הוא סטטיסטי ולא מוחלט: שושלות בודדות עשויות לרדת, אך הממוצע האוכלוסייתי משתפר. קיים חץ כיווני לעבר כיסוי רגולטורי מקיף יותר והולך.
הערת תורה (הצעה תיאורטית, לא הוכחה)
הפסוק "לֹא טוֹב הֱיוֹת הָאָדָם לְבַדּוֹ" (בראשית ב:יח) — ללא נקבה: אין piRNA ואין לחץ ברירה. "אֶעֱשֶׂה לּוֹ עֵזֶר כְּנֶגְדּוֹ" — עֵזֶר = מסייע (piRNA מסייע לרגולציה), כְּנֶגְדּוֹ = כנגד (דוחה זכרים בלתי מרוסנים רגולטורית). "וְהָאָדָם יָדַע אֶת חַוָּה" (בראשית ד:א) — ידע/דעת מתאימים לתדירות של 33%F, הפועל הרגולטורי. "אֶרֶץ... תֹּלֶה אֶרֶץ עַל בְּלִימָה" (איוב כו:ז) — הגנום מוחזק על ידי ריסון (piRNA אימהי). הנקבה היא המנוע הרגולטורי: היא מספקת הן הוראות (piRNA) והן לחץ ברירה (בחירת בן זוג).
מה טענה זו אינה גורסת
אין אנו טוענים כי ברירה מינית "תוכננה" באופן תכליתי. הטענה היא כי בחירת בן זוג על בסיס סימטריה מהווה, מבחינה מכניסטית, ברירה לשלמות רגולטורית של אלמנטים ניידים. ההקבלות התורניות מוצעות בהתאם למתודולוגיה המפורטת בסעיף 18.
סיכום: ברירה מינית לסימטריה = ברירה לשלמות רגולטורית. הנקבה מניעה את התהליך באמצעות בחירת בן זוג ותרומת piRNA. הזכר תורם חלבוני KRAB-ZFP. יחד הם מייצרים חץ כיווני לעבר רגולציה משופרת. מושגי התורה "עזר כנגדו" ו"ידע" מתיישבים מבנית עם מודל זה.
9. גני גדילה: מוגנים עד שנפרצים
העיקרון הוויסותי פועל לא רק באמצעות מה שמופעל, אלא באמצעות מה שמוגן. גני ההתפתחות הקריטיים ביותר נשמרים באופן שיטתי מפני חדירת טרנספוזונים.
| גן | תפקוד | %TE | מול הגנום (45.7%) |
|---|---|---|---|
| IGF2 | גורם גדילה (מוטבע) | 17.1% | ×0.37 |
| FGFR3 | קולטן גדילה | 22.3% | ×0.49 |
| GH1 | הורמון גדילה | 30.1% | ×0.66 |
| IGF1R | קולטן IGF1 | 31.7% | ×0.69 |
| IGF1 | גורם גדילה | 38.9% | ×0.85 |
הדפוס משקף את SHH בבקר (×0.45): וויסותנים התפתחותיים קריטיים מוגנים מפני חדירת טרנספוזונים. ענקיוּת (gigantism) היא פריצה של ההגנה — טרנספוזון בלוקוס גדילה שבדרך כלל נשמר נקי.
אותו דפוס הגנה מופיע ב-SHH בבקר: ×0.45, בהתאמה ל-IGF2 כגן הדל ביותר בטרנספוזונים בגנום שלו. הוויסותנים ההתפתחותיים הקריטיים — גדילה, דפוסיות (patterning), הטבעה גנומית (imprinting) — מוגנים באופן אוניברסלי.
המקבילה הקלינית היא תסמונת Beckwith-Wiedemann: אובדן הטבעה של IGF2 → ביטוי-יתר של IGF2 → גדילת-יתר עוברית. המנגנון הוא בדיוק מה שדפוס דלדול ה-TE מנבא: פרצו את אזור ההגנה סביב IGF2, והגדילה חומקת מוויסות.
תיאור התורה על ענקים שלאחר המבול ("הנפילים היו בארץ בימים ההם וגם אחרי כן" — בראשית ו:ד) מתאר פנוטיפ העולה בקנה אחד עם דיסרגולציה ספורדית של גדילה בתיווך טרנספוזונים, שחוזרת ומופיעה ("וגם אחרי כן") מווריאנטים סמויים באוכלוסייה שלאחר צוואר-בקבוק. הענקים מוזכרים בשמם הפרטי — עוג מלך הבשן, שלושת בני ענק בחברון, הרפאים, האימים — משום שכל אחד מייצג פריצה וויסותית נדירה, לא אוכלוסייה. ראוי לציון שענק יושב בחברון, בנחלת כנען בן חם — השושלת שהונחה עליה קללה (בראשית ט:כה). אם לקללה יש מתאמים גנומיים, דיסרגולציית גדילה בתיווך טרנספוזונים בשושלת פטרילינאלית ספציפית היא אפשרות ביולוגית סבירה.
סיכום: גני גדילה (IGF2, GH1, IGF1) דלים בטרנספוזונים — מוגנים מפני חדירה, כשם ש-SHH מוגן בבקר. ענקיוּת = פריצה של הגנה זו. התורה קוראת לכל ענק בשמו משום שכל אחד מייצג כשל וויסותי נדיר, לא אוכלוסייה.
10. הבסיס הזוחלי: כאוס, יציבות וחזרה
עד כה בחנו את העיקרון הוויסותי במעלי-גרה (שיווי-משקל BovB/L1), בפרימטים (הפעלת L1HS) ובגני התפתחות (דלדול TE). אותו עיקרון מארגן את ההתבדלות העמוקה ביותר באבולוציית בעלי-החוליות: הזוחלים וצאצאיהם.
| מין | גנום | TE כולל | DNA-TEs | פרשנות |
|---|---|---|---|---|
| תנין (Alligator) | 2.18 Gb | 37.7% | **18.0%** | כאוטי — "גזור והדבק" פעיל |
| צב (Turtle) | 2.13 Gb | 15.0% | 3.3% | יציב — ללא שינוי 200 מיליון שנה |
| תרנגולת (Chicken) | 1.05 Gb | 12.8% | 1.0% | קומפקטי — יורשת הדינוזאורים, חזרה |
| בקר (Cattle) | 2.67 Gb | 50.5% | 2.3% | מאורגן — שיווי-משקל BovB/L1 |
התנין — קרוב המשפחה החי הקרוב ביותר לדינוזאורים — מכיל 18% טרנספוזונים מסוג DNA, הצורה המשבשת ביותר של כאוס גנומי. התרנגולת — צאצא ישיר של הדינוזאורים — דחסה את הגנום שלה למחצית הגודל היונקי. יורשת הדינוזאורים לא המשיכה להתנפח. היא חזרה לוויסות.
כפי שהוכח בסעיף 1, הנחש העביר את BovB ליונקים בהעברה אופקית (Walsh et al. 2013; Ivancevic et al. 2018). גיל BovB בבקר (~22 מיליון שנה לפי שעון ההתבדלות) צעיר יחסית ל-L1 (~43 מיליון שנה) — הגעה זרה ומאוחרת שהוטמעה בארכיטקטורה הוויסותית של הפונדקאי. ההבטחה לאישה — "הרבה ארבה עצבונך והרונך" (בראשית ג:טז) — עולה בקנה אחד עם ההעשרה של BovB בגנים הקשורים לרבייה ולנפש (חיוּת פיזית): MHC, קולטנים אולפקטוריים ולוקוסים של רבייה — כולם מועשרים ב-BovB במעלי-גרה.
התנין, עם 65 משפחות TE שונות ו-18% טרנספוזונים מסוג DNA (מחלקת ה"גזור והדבק" — הצורה המשבשת ביותר של פעילות טרנספוזונים), מייצג כאוס גנומי. הצב, שלא השתנה במשך 200 מיליון שנה עם 15% TE בלבד ומשפחה דומיננטית אחת (CR1, בשיעור 54%), מייצג יציבות קדומה. התרנגולת — צאצא ישיר של דינוזאורים תרופודיים שייצרו פעם את בעלי-החיים היבשתיים הגדולים ביותר בתולדות כדור הארץ — דחסה את הגנום שלה ל-1.05 Gb (מחצית מהממוצע היונקי), עם 12.8% TE בלבד. יורשת הדינוזאורים חזרה לוויסות.
אלה אינן אבסטרקציות. התנין הוא קרוב המשפחה החי הקרוב ביותר לאורגניזמים ששלטו בכדור הארץ במשך 165 מיליון שנה. הגנום שלו נקרא כמו אתר ארכאולוגי: שכבה על שכבה של פריצות טרנספוזונים בלתי-תלויות, אף אחת מאורגנת, אף אחת בשיווי-משקל. הגנום של הבקר, לעומת זאת, ב-50.5% TE — יותר תוכן טרנספוזוני מהתנין — הוא מובנה: אלמנטים מסוג LINE (L1 ו-BovB) מהווים 28.3%, מאורגנים לשתי מערכות מאוזנות. תוכן גבוה, אך מווסת. העיקרון המארגן אינו "כמה" אלא "כיצד מנוהל".
סיכום: התנין (37.7% TE, 18% טרנספוזוני DNA) = כאוס גנומי. הצב (15% TE, ללא שינוי 200 מיליון שנה) = יציבות קדומה. התרנגולת (1.05 Gb, יורשת דינוזאורים) = חזרה לוויסות. הנחש העביר BovB ליונקים (הגברה ×2,151) אך כמעט לא שמר — "ארור מכל הבהמה." וויסות, לא מורכבות, הוא המבחין בין מצבים גנומיים.
11. שני שבועות, שני גנומים: המעבר הזוחלי
המקרא העברי נפתח בשני סיפורי בריאה. הם מפעילים שמות אלוהיים שונים, פעלים שונים, מונחי בעלי־חיים שונים — והבחנות אלו ממופות על ההבדל שבין גנומים זוחליים כאוטיים מבחינת אלמנטים טרנספוזביליים (TE) לבין גנומים עופיים ויונקיים מווסתים מבחינת TE.
שני סיפורי בריאה, שני שמות אלוהיים
בראשית א (שבוע ראשון) = באופן בלעדי אלהים. יום חמישי: "וַיִּבְרָא אֱלֹהִים אֶת הַתַּנִּינִם הַגְּדֹלִים... וְאֵת כָּל עוֹף כָּנָף" (בראשית א:כא). יום שישי: "תּוֹצֵא הָאָרֶץ נֶפֶשׁ חַיָּה... בְּהֵמָה וָרֶמֶשׂ וְחַיְתוֹ אָרֶץ" (בראשית א:כד). מונחים מכריעים: תנינם הגדלים (הזוחלים הגדולים), עוף כנף (עוף בעל כנף — אבר פיזי), חיתו ארץ (חיות הארץ — חומר גולמי). הפועל: ברא (יצירה יש מאין). הפועל: אלהים בלבד.
בראשית ב (שבוע שני) = יהוה אלהים (שני השמות יחדיו). "וַיִּצֶר יְהוָה אֱלֹהִים מִן הָאֲדָמָה כָּל חַיַּת הַשָּׂדֶה וְאֵת כָּל עוֹף הַשָּׁמָיִם" (בראשית ב:יט). הפועל: יצר (עיצוב בכוונה, פועל היוצר — כיוצר חרס). בעלי החיים מכונים מחדש: עוף השמים (עוף השמים — תחום), חית השדה (חיית השדה — מרחב מווסת). תנינם הגדלים נעדרים לחלוטין מן השבוע השני.
ההבחנה הלשונית
כנף: כ(BKL) + נ(AMTN) + ף(F) = אבר פיזי המכה או מכסה, כנף כמכשיר.
שמים: ש(F) + מ(AMTN) + י(YHW) + ם(AMTN) = 25%F, תחום שמימי. מגדיר את העוף על פי הֵיכָן הוא שייך, לא על פי מָה יש לו.
ארץ = אדמה פיזית, חומר.
שדה = שדה, מרחב מעובד, קרוב מבחינה אטימולוגית ל־שדי (אל שדי) — התחום המווסת.
תנינם הגדלים = רק בשבוע הראשון. לעולם לא בשבוע השני. יצורים אלו שייכים באופן בלעדי למשטר של אלהים בלבד.
המיפוי הגנומי
שבוע ראשון (אלהים) = כאוס של TE: באליגטור 37.7% TE, 18% DNA-TEs, 65 משפחות. בדינוזאורים הערכה של 2–3 Gb (Organ et al. 2007). אין שיווי־משקל BovB/L1. ריבוי טהור. אלהים = השם של "פְּרוּ וּרְבוּ" (ריבוי), בריאה, חוק הטבע.
שבוע שני (יהוה אלהים) = מווסת: עופות מודרניים 1.05 Gb, 12.8% TE. מעלי גרה עם BovB/L1. יונקים עם piRNA, KRAB-ZFP. הוספת יהוה = הוספת וויסות מכוון.
הנחש מכיר רק שם אחד
בראשית ג:א: הנחש אומר "אַף כִּי אָמַר אֱלֹהִים" — רק אלהים, לעולם לא יהוה. שייך למשטר שקדם לוויסות. BovB = 0.01% = נתן הכול, לא שמר מאומה.
הזמן בשבוע הראשון
אין שמש עד יום רביעי (בראשית א:יד–טז). "יְהִי מְאֹרֹת... לְאֹתֹת וּלְמוֹעֲדִים וּלְיָמִים וְשָׁנִים" — זמן מדיד מתחיל ביום הרביעי. לפני כן: משטר אלהים, משך בלתי ידוע. עידן המזוזואיקון משתלב כאן באופן טבעי.
המבול כגבול משטרי
מה שנכנס לתיבה = שורד אל תוך משטר יהוה אלהים. מחוצה לה = נכחד. 8 בני אדם = צוואר בקבוק → קריסת piRNA → נפילת תוחלת חיים פי 35.6.
מה טענה זו אינה גורסת
אין כאן טענה שהפליאונטולוגיה טועה. בין אם העופות התפתחו מתרופודים ובין אם נבראו בנפרד, המעבר הגנומי זהה: כאוס TE → וויסות. מסגרת שני השבועות = קריאה מבנית הממופה על הנתונים.
סיכום: בראשית א (אלהים) = ריבוי ללא וויסות מכוון — בהתאמה לכאוס TE זוחלי. בראשית ב (יהוה אלהים) = וויסות מכוון — בהתאמה לגנומים עופיים קומפקטיים וגנומים יונקיים מווסתים. המונחים מתחלפים בדיוק: כנף→שמים, ארץ→שדה. הנחש מכיר רק את אלהים. המבול = גבול משטרי.
12. משוואת חומר/רוח
הנתונים שהוצגו עד כה — יחסי BovB/L1, הפעלת L1HS, דלדול TE, קווי-בסיס זוחליים — מתארים מערכת פיזית. אולם התורה מקודדת את אותן הבחנות עצמן במורפולוגיה שלה. ניתוח אחוז היסוד (F%), שאומת על-פני 98,122 זוגות מילים בדיוק ניבויי של 87.8%, מייחס כל אות לקבוצה תפקודית. הגרדיאנט הנוצר ממופה ישירות על הנתונים הגנומיים.
אחוז היסוד (F%) מייחס כל אות עברית לאחת מארבע קבוצות תפקודיות. ביישומו על שמות המקודדים את הקוטביוּת חומר–רוח:
| F% | קטגוריה | דוגמאות | מקבילה גנומית |
|---|---|---|---|
| 0% | וויסות טהור | יהוה, אלהים | מערכת ניהול L1HS |
| 25% | הפעלה | נשמה, אד-ני | L1HS עצמו — המתג |
| 40% | אדם מרומם | ישראל | L1HS מווסת + תורה |
| 50% | אדם בסיסי | יעקב | L1HS פעיל, לפני התרוממות |
| 67% | כבד-חומר | עשו, עפר | פנוטיפ של קוף: L1HS כבוי |
| 75% | עודף חומר | שעיר | ביטוי-יתר של KRTAP |
| 100% | חומר טהור | שער (שיער) | קרטין — הרקמה העשירה ביותר ב-BovB |
שינוי השם מיעקב (50%) לישראל (40%) מקודד שדרוג וויסותי — פחות חומר, יותר שליטה. המילה אדמה (25% F) מקבלת אות אחת נוספת והופכת לאדם — ואותה אות היא ה, אות הבדלה מקבוצת יה"ו. במערכת המורפולוגית שתיעדנו על-פני 98,122 זוגות מילים, אותיות יה"ו הן המנגנון המדויק שבו שורשים מתבדלים למשמעויות שונות.
הגנום שכל היונקים חולקים הוא האדמה. השכבה שהופכת מין אחד לאנושי היא הה.
סיכום: גרדיאנט ה-F% — משער (100%, חומר טהור) דרך עשו (67%) ויעקב (50%) ועד יהוה (0%, וויסות טהור) — ממופה על הספקטרום הגנומי מרקמה הנשלטת על-ידי BovB (קרטין) ועד קוגניציה המופעלת על-ידי L1HS. המילה אדם נבדלת מאדמה באות הבדלה אחת מקבוצת יה"ו — אותה מחלקת אותיות שמבדילה משמעויות על-פני כל 98,122 זוגות המילים שנבדקו.
13. גיוון מהיר: מנוע ה-BovB
הפרקים הקודמים ביססו את הארכיטקטורה הרגולטורית כעקרון המארגן של מינים קיימים. שאלה טבעית עולה בעקבותיהם: האם אותו מנגנון יכול להסביר גם את מקורו של מגוון המינים?
היתכנות של גיוון מהיר ממאגר אבות מצומצם:
תקדים. דגי ציקליד באגם ויקטוריה: כ-500 מינים מאב קדמון אחד בתוך כ-15,000 שנה (Seehausen 2006). ממצא שפורסם בביקורת עמיתים, בלתי שנוי במחלוקת.
מנגנון. BovB בלוקוסים שונים → פנוטיפים שונים. BovB בלוקוס KRTAP → קרניים קרטיניות (Bovidae). BovB בלוקוס SHH → ניבים (Moschidae). אותו טרנספוזון, מטרה שונה, מין שונה. אתרי השתלבות: 94.5% משותפים בין פרה↔כבש (±10 kb), 74% בין פרה↔אייל, 51.3% בין כבש↔אייל — מפל פילוגנטי ממוצא משותף.
קצב. BovB עדיין פעיל בבקר: 7,325 עותקים בסטייה של פחות מ-2% מרצף הקונצנזוס, המעידים על טרנספוזיציה מתמשכת. בכבשים נמצאו 4,854 עותקים צעירים; באיילים רק 427 — מפל של ירידה בפעילות BovB, המשקף את יחס BovB/L1 עצמו. בקירוב של השתלבות חדשה אחת לדור לאורך כ-1,667 דורות ב-5,000 שנה, שושלת המעלי גרה הייתה חשופה לכ-133,000 הזדמנויות השתלבות. כדי לייצר 200 מינים מ-10 טיפוסים קדומים נדרש "שיעור הצלחה" של 0.15% בלבד.
עדות קריוטיפית. המונטג׳ק מספק הוכחה חיה לשינוי כרומוזומלי מונע טרנספוזונים: ל-Muntiacus muntjak יש 2n=6 (מספר הכרומוזומים הנמוך ביותר בכל היונקים), בעוד שלמונטג׳ק ריבס יש 2n=46 — בתוך סוג (genus) אחד. אלה אינם שינויים קדומים. הם מוכיחים שמיזוג ופיצול כרומוזומלי בתיווך טרנספוזונים יכולים לשנות מבנה גנום באופן קיצוני בסולמות זמן אבולוציוניים קצרים. Fedoroff (2012, Science) ו-Chuong et al. (2017, Nature Reviews Genetics) טענו באופן בלתי תלוי כי יסודות נודדים משמשים כמנועי מינוּת (speciation engines), המספקים את חומר הגלם לבידוד רבייתי ולחדשנות רגולטורית.
סיכום: גיוון מהיר ממאגר אבות מצומצם הוא אפשרי מבחינה ביולוגית. דגי ציקליד ייצרו 500 מינים ב-15,000 שנה. BovB משתלב בלוקוסים שונים כדי לייצר מורפולוגיות שונות מאותו גנום. BovB עדיין פעיל בבקר (7,325 עותקים צעירים). המונטג׳ק מדגים שינוי קריוטיפי מונע טרנספוזונים בתוך סוג אחד (2n=6 עד 2n=46).
14. שלוש מדרגות רגולציה
הנתונים משתרעים כעת על פני מעלי גרה, פרימטים, זוחלים, גנים של גדילה ומנגנוני מינוּת. דפוס עולה שאינו נראה מתוך אף מערך נתונים בודד, אך מתבהר כשקוראים אותם יחד: הרגולציה הביולוגית פועלת ברמות היררכיות, כל אחת בנויה על קודמתה.
הנתונים חושפים שלוש רמות היררכיות של רגולציה ביולוגית, כל אחת בנויה על קודמתה:
רמה 1: בהמה — רגולציה אוטומטית. שיווי המשקל BovB/L1 במעלי גרה פועל ללא קוגניציה. היחס מתקיים בכוח מנגנונים מולקולריים — ברירה, מתילציה, העדפת השתלבות. המילה העברית "בהמה" ניתנת לקריאה כ-ב-ה-מ-ה: "בה מה" — בה הכוחות. זוהי רגולציה כהומאוסטזיס. הכבש ביחס 1.00 הוא ביטויה הטהור ביותר.
רמה 2: אדם — רגולציה מודעת. L1HS פעיל בנוירונים אנושיים — ובפרט בהיפוקמפוס, מושב הלמידה והזיכרון. Coufal et al. (2009) הדגימו רטרוטרנספוזיציה של L1 בתאי אב עצביים (neural progenitor cells). כל השתלבות חדשה משנה את ביטוי הגנים באותו נוירון ויוצרת מצבים רגולטוריים ייחודיים. זו אינה הומאוסטזיס. זוהי הסתגלות. למידה. בחירה. הרגולציה אינה עוד אוטומטית — היא מנוהלת על ידי התודעה.
רמה 3: תורה — רגולציה מכוּונת. אם L1HS מספק את המנגנון לנוירופלסטיות, ונוירופלסטיות מאפשרת למידה, אזי מערכת של למידה מכוּונת — תרגול התנהגותי מובנה — מהווה שכבה רגולטורית שלישית. התורה (מן השורש הו"ר, "הוראה") מספקת 613 פרוטוקולים התנהגותיים. כל שינוי התנהגותי מתפשט דרך מסלולים מתועדים היטב: התנהגות → פעילות עצבית → שינוי אפיגנטי → שינוי בביטוי גנים. התורה אינה מתארת רגולציה. התורה היא רגולציה — שכבת קושחה (firmware) עבור המערכת הרגולטורית המודעת ש-L1HS מאפשר.
היררכיה זו מיישבת שאלה שהנתונים מעלים אך אינם יכולים לענות עליה בכוחות עצמם: מדוע L1HS פעיל בבני אדם ולא בקופות? התשובה המולקולרית (הגנה חזקה יותר של APOBEC3B/PIWIL2) מסבירה כיצד. ההיררכיה הרגולטורית מציעה מדוע: L1HS דורש מערכת ניהול. בבעל החיים, BovB/L1 הוא מנוהל עצמית. באדם, L1HS דורש ניהול מודע. בלי מערכת של תרגול מכוּון — בלי "הוראות" — המתג מסוכן (תסמונת Rett) במקום פרודוקטיבי.
המסלול המולקולרי מהתנהגות לביטוי גנים מתועד היטב: מעורבות התנהגותית → ירי עצבי → הפעלת גנים מיידיים מוקדמים (cFos, BDNF) → אצטילציה של היסטונים → שיפוץ כרומטין (chromatin remodeling) → דה-מתילציה של L1 → השתלבות L1 חדשה → שינוי בביטוי גנים באותו נוירון. Maze et al. (2011) הדגימו מסלול זה ישירות: חשיפה לקוקאין הפעילה רטרוטרנספוזיציה של L1 בגרעין המאסף (nucleus accumbens) ושינתה את מעגלי התגמול. המסלול הוא ממשי, פורסם, ואינו ייחודי לפתולוגיה — הוא פועל בכל מקרה שבו מעורבות עצבית מתמשכת מייצרת שינוי אפיגנטי.
לימוד תורה, במסגרת מודל זה, הוא מעורבות עצבית מובנית החוצה את כל התחומים ההתנהגותיים. 613 המצוות מקיפות תזונה (כשרות), שינה (קריאת שמע על המיטה), דיבור (הלכות לשון הרע), עבודה (מנוחת שבת), רבייה (טהרת המשפחה), חקלאות (שמיטה) ומשפט (בתי דין). אין תחום התנהגותי שאינו מוסדר. כל פרוטוקול התנהגותי, כשהוא מתורגל באופן עקבי, מפעיל מעגלים עצביים ספציפיים, מייצר שינויים אפיגנטיים ספציפיים, ואם מנגנון L1HS פועל כפי שתועד — מייצר מצבים רגולטוריים ספציפיים בנוירונים ספציפיים. התורה אינה תיאור של רגולציה. היא פרוטוקול התנהגותי שדרך מסלולים מולקולריים מתועדים מייצר רגולציה.
סיכום: שלוש רמות רגולציה — אוטומטית (בהמה: שיווי משקל BovB/L1), מודעת (אדם: L1HS בנוירונים), ומכוּונת (תורה: פרוטוקולים התנהגותיים המייצרים שינוי אפיגנטי דרך מסלולים מולקולריים מתועדים). התורה אינה מתארת רגולציה. היא רגולציה — שכבת קושחה עבור המערכת המודעת ש-L1HS מאפשר.
15. בראשית ג׳: האירוע הרגולטורי
סיפור התורה על גן העדן, הנקרא דרך המסגרת הרגולטורית שהוצגה בפרק זה, מתאר מעבר גנומי ספציפי: מוויסות אוטומטי לוויסות מודע, שנגרם על ידי החדרת אלמנט גנטי זר.
הסוכן: חומר טהור
אחוז היסוד של הסוכנים המרכזיים בסיפור מעורר תשומת לב:
| מילה | אותיות | %F | קטגוריה |
|---|---|---|---|
| נחש (serpent) | נ(A)+ח(F)+ש(F) | 67% | כבד-חומר |
| זרע (seed) | ז(F)+ר(F)+ע(F) | 100% | **חומר טהור** |
המילה זרע — זרע, צאצא, חומר גנטי — היא 100% יסוד. כל אות בה היא אות תוכן. בשפה שבה השמות האלוהיים מכילים 0% יסוד, המילה המציינת את התרומה הגנטית של הנחש היא חומר פיזי טהור.
לעומת זאת, אוצר המילים שלאחר האירוע משתנה באופן דרמטי:
| מילה | %F | קטגוריה |
|---|---|---|
| דעת (knowledge) | 33% | רגולטורית |
| מות (death) | 0% | רגולטורית טהורה |
| איבה (enmity) | 0% | רגולטורית טהורה |
| כתנת (garment) | 0% | רגולטורית טהורה |
המילים שלפני האירוע (טוב, חיים, ערום) מציגות ממוצע של 42.9% יסוד. המילים שלאחר האירוע (דעת, מות, איבה, כתנת, עור) מציגות ממוצע של 33.3% — ירידה של 9.5 נקודות אחוז. הסיפור נע לעבר תוכן רגולטורי רב יותר, לא פחות. "דעת טוב ורע" היא, מבחינה מורפולוגית, עלייה בכושר הרגולטורי.
השומרים: גני הגנה מדוללי-TE
בראשית ג׳:כ״ד מתאר את מה שהוצב לשמור על הנתיב חזרה למצב המקורי:
"ויגרש את האדם וישכן מקדם לגן עדן את הכרבים ואת להט החרב המתהפכת לשמר את דרך עץ החיים"
הגנום מכיל מערכת הגנה כנגד רטרוטרנספוזיציה של L1 — אותו אלמנט עצמו שהפעלתו מבדילה בין בני אדם לקופים. מדדנו את צפיפות ה-TE בגנים המקודדים להגנה זו:
| גן | תפקיד | TE × גנום | סטטוס |
|---|---|---|---|
| PIWIL1 | מסלול piRNA (השתקת L1) | **×0.56** | **מוגן** |
| DNMT3A | מתילציה de novo | **×0.72** | **מוגן** |
| PIWIL2 | מסלול piRNA | ×0.89 | מדולל במידה קלה |
| DNMT3B | מתילציה de novo | ×0.86 | מדולל במידה קלה |
| APOBEC3B | הרס RNA של L1 | ×1.16 | לא מדולל |
| PIWIL4 | מסלול piRNA | **×1.88** | **נפלש על ידי L1** |
PIWIL1, גן ההגנה העיקרי של מסלול ה-piRNA, מדולל ב-TE ברמה של ×0.56 — המתקרבת לרמת ההגנה של IGF2 (×0.37), הגן המוגן ביותר בגנום. DNMT3A, ה-methyltransferase של מתילציה de novo הכותב את סימני המתיל המשתיקים את L1, מוגן ברמה של ×0.72. אלה הם "כרובי" הגנום — שומרים סטטיים, מדוללי-TE, המקיימים את המחסום בין L1 פעיל לבין המצב האוטו-רגולטורי.
APOBEC3B פועל באופן שונה: הוא אנזים פעיל המבצע דה-אמינציה של RNA של L1 ומשמיד אותו כימית. הוא אינו מדולל-TE (×1.16) מפני שאינו זקוק להגנה פסיבית — הוא עצמו נשק. "חרב להט."
המילה מתהפכת מוסיפה דיוק: הגנתו של APOBEC3B אינה קבועה. היא מתאפסת בכל דור כאשר המתילציה עוברת תכנות מחדש בקו הנבט. בכל דור, החרב "מתהפכת" — ההגנה חייבת להיות מוקמת מחדש. Baduel et al. (2025) תיעדו איפוס בין-דורי זה ביונקים.
ו-PIWIL4 — גן piRNA שתפקידו להשתיק את L1 — נפלש על ידי L1 ברמה של ×1.88. הנחש נמצא בתוך השומר. מירוץ החימוש שתיארו Jacobs et al. (2014) אינו מטאפורה המוחלת על בראשית. הוא מציאות נמדדת שאליה בראשית ממופה.
שני אירועים, שני יסודות
הסיפור מבחין בין שני אירועים:
1. תרומת הנחש — "בין זרעך ובין זרעה" (בראשית ג׳:ט״ו). BovB: רטרוטרנספוזון זוחלי שהועבר אופקית, כפי שתועד על ידי Walsh et al. (2013). "זרע הנחש" הוא, מבחינה גנומית, מציאות נמדדת.
2. תוצאת העץ — "דעת טוב ורע" (בראשית ג׳). רכישת כושר רגולטורי: היכולת להבחין, לנהל, לבחור. בגנום, הדבר ממופה למערכת הניהול של L1HS — APOBEC3B, PIWIL2, מנגנון המתילציה — המאפשרת פעילות מבוקרת של L1 במוח האנושי.
לפני האירוע: וויסות אוטומטי. "עץ החיים" מייצג מצב שבו BovB נעדר ו-L1 אינו דורש ניהול מודע. זוהי רמה 1: בהמה.
לאחר האירוע: נדרש וויסות מודע. BovB נוכח כעת בגנום (זרע הנחש). L1 אינו יכול עוד להישאר על טייס אוטומטי. על האדם לפתח ולקיים מערכת ניהול — או לשאת בתוצאות (תסמונת Rett: כשל MeCP2 → L1 בלתי מבוקר → קריסה נוירולוגית).
"כתנות עור" (בראשית ג׳:כ״א) הן הדבר הראשון שניתן לאחר המעבר הרגולטורי. עור הוא הרקמה בעלת העשרת BovB הגבוהה ביותר בגוף היונק: KRTAP ב-22.5%. הלבוש אינו מטאפורה לצניעות. הוא הרקמה הראשונה העשירה ב-BovB — הסימן הפיזי של המצב הגנומי החדש.
עלה התאנה: הכיסוי הראשון
בראשית ג׳:ז׳ מתעד את התגובה הראשונה לרכישת הדעת: "ויתפרו עלה תאנה" — "הם תפרו עלי תאנה." הבחירה בתאנה אינה מקרית.
התאנה (תאנה: ת-א-נ-ה) היא 0% יסוד — אותיות רגולטוריות טהורות, ללא תוכן פיזי. היא חולקת תכונה זו עם השמות האלוהיים יהוה ואלהים. הצרעה (צרעה: צ-ר-ע-ה) המאביקה אותה היא 75% יסוד — כבדת-חומר, והיא מכילה בתוך אותיותיה את המילה רע (100% יסוד).
התאנה אינה יכולה להתרבות ללא הצרעה. הצרעה מתה בתוך התאנה. זו אינה מטאפורה — זהו מוטואליזם אובליגטורי מתועד. צרעת התאנה (Blastophaga psenes) חודרת לתאנה, מאביקה אותה ומתה בתוכה. ללא מוות זה, התאנה אינה מניבה פרי. ה"רע" (רע, 100% חומר) חייב להיכנס לתוך ה"טהור" (תאנה, 0% חומר) ולהיצרך כדי שהחיים יימשכו.
הכיסוי הראשון הוא רגולטורי (עלה תאנה, 0% F). הכיסוי השני, שניתן על ידי אלוהים, הוא פיזי: כתנות עור (בראשית ג׳:כ״א). עור הוא הרקמה בעלת העשרת BovB הגבוהה ביותר: KRTAP ב-22.5%. הסיפור נע מתגובה רגולטורית למציאות פיזית — מ-0% יסוד אל הרקמה העשירה ביותר ב-BovB בגוף היונק.
הווירוס שהפך לאמהות
הדפוס שבו יסודות זרים הופכים חיוניים לחיים חורג מגבולות הטרנספוזונים. Syncytin-1, החלבון החיוני ליצירת שכבת ה-syncytiotrophoblast בשליית האדם, מקודד על ידי גן המעטפת של רטרווירוס אנדוגני (HERV-W). Mi et al. (2000, Nature 403:785) הדגימו כי syncytin מתווך מיזוג תא-תא בשלייה. בלעדיו אין שלייה נוצרת; בלא שלייה — אין רבייה של יונקים.
Syncytin נלכד באופן בלתי תלוי לפחות שש פעמים בשושלות יונקים שונות (Lavialle et al. 2013) — רטרווירוסים שונים, אותו תפקיד חיוני, ביות מתכנס. "טפיל" הפך לתנאי-שאין-בלתו של אמהות.
בראשית ג:כ: "וַיִּקְרָא הָאָדָם שֵׁם אִשְׁתּוֹ חַוָּה כִּי הִוא הָיְתָה אֵם כָּל חָי." להיות אם כל חי, ביונקים, מחייב שלייה. לקיים שלייה מחייב רטרווירוס. קריאת השם מתרחשת מיד לאחר הקללות, כאילו מתוך הכרה: המציאות הרבייתית החדשה — התלויה בגן ויראלי — היא מעתה הדרך קדימה.
Syncytin אינו מקרה בודד. כ-5–8% מהגנום האנושי מורכב מרצפים רטרו-ויראליים אנדוגניים — פי שלושה עד חמישה יותר DNA ויראלי מ-DNA מקודד-חלבון. בין הביותים המתועדים: HERV-K פעיל החל משלב העובר בן שמונה התאים, וחלבון ה-Rec שלו מספק הגנה אנטי-ויראלית (Grow et al. 2015, Nature). אלמנטים מסוג MER41 משמשים כמגבירים הניתנים להשראה על ידי אינטרפרון; מחיקתם באמצעות CRISPR גורמת לקריסת התגובה החיסונית המולדת (Chuong, Elde & Feschotte 2016, Science). תשעים אחוז משרידי הרטרווירוסים האנדוגניים הם LTRs בודדים המתפקדים כמקדמים ומגבירים, כאשר 64% מהתמלילים המונעים על ידי LTR מבוטאים ברקמות רבייה. הגנום האנושי אינו רק "מזוהם" על ידי וירוסים. הוא מופעל על ידם.
התמוטטות אורך החיים
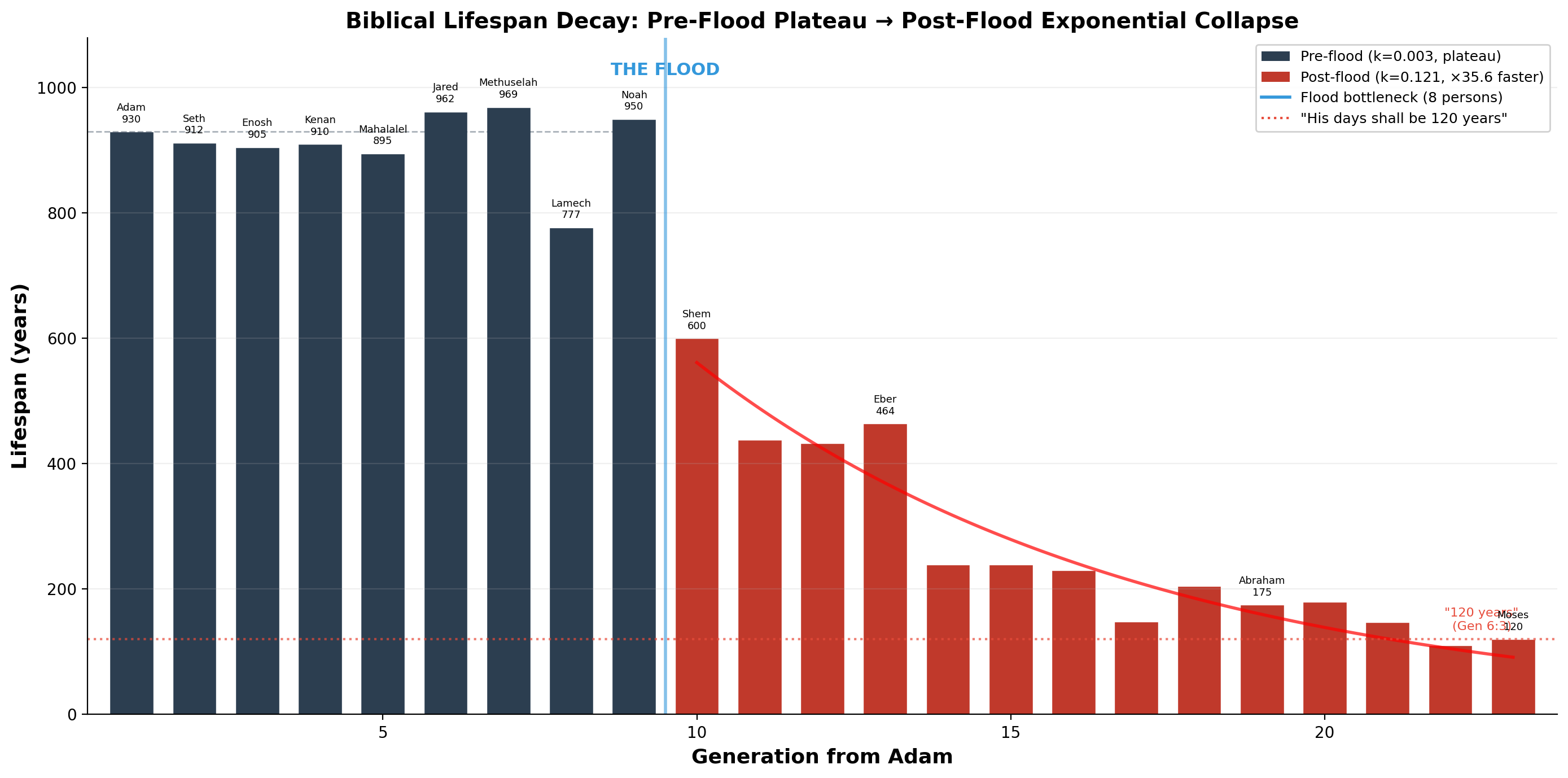
התורה מתעדת ירידה מדויקת באורך חיי האדם לאורך הדורות:
| דור | שם | אורך חיים | שלב |
|---|---|---|---|
| 1 | אדם | 930 | רמה טרום-מבולית |
| 7 | מתושלח | 969 | רמה טרום-מבולית |
| 10 | נח | 950 | אחרון טרום-מבולי |
| 11 | שם | 600 | קריסה בתר-מבולית |
| 14 | עבר | 464 | |
| 16 | פלג | 239 | |
| 20 | אברהם | 175 | |
| 22 | יעקב | 147 | |
| 26 | משה | 120 | גבול אסימפטוטי |
התאמת מודל דעיכה מעריכית חושפת שני שלבים מובחנים:
- טרום-מבול: קבוע דעיכה k = 0.0034 (רמה כמעט שטוחה בסביבות 930 שנה)
- בתר-מבול: קבוע דעיכה k = 0.1207 (מהיר פי 35.6 מהשלב הטרום-מבולי)
הקריסה אינה הדרגתית. היא פתאומית — מתחילה מיד לאחר צוואר הבקבוק של המבול, כאשר האוכלוסייה האנושית התכווצה לשמונה פרטים.
De Cecco et al. (2019, Nature) הדגימו כי רטרוטרנספוזונים מסוג L1 מופעלים במהלך ההזדקנות בתאים סומטיים, מפעילים את מסלול החיסון המולד cGAS-STING ומייצרים דלקת כרונית — החתימה המולקולרית של ההזדקנות. הפעלת L1 אינה תוצאה של הזדקנות; היא מנוע ההזדקנות.
צוואר בקבוק אוכלוסייתי של שמונה פרטים היה מצמצם באופן קטסטרופלי את מגוון רצפי ה-piRNA, וריאנטים של KRAB-ZFP ודפוסי המתילציה המשתיקים יחד אלמנטים ניידים. כל דור עוקב, עם מגוון השתקה מצומצם יותר, היה חווה שחרור-דיכוי גובר של אלמנטים ניידים — מצטבר, מתגבר, מעריכי.
בראשית ו:ג: "וְהָיוּ יָמָיו מֵאָה וְעֶשְׂרִים שָׁנָה." עקומת הדעיכה המעריכית שואפת לערך של כ-120 — הגבול שנקבע עוד לפני שהמבול אירע. משה חי בדיוק 120 שנה (דברים לד:ז).
סיכום: בראשית ג מתאר מעבר מוויסות אוטומטי לוויסות מודע, שנגרם על ידי החדרת חומר גנטי זר (נחש ← BovB). אחוז ה-F עובר ממשקל-חומרי (סוכנים: 83%) לוויסותי (תוצאות: 33%). גני הגנה (PIWIL1 ×0.56) מוגנים; אחרים (PIWIL4 ×1.88) נפלשים. עלה התאנה (0%F) הוא התגובה הוויסותית הראשונה; העור (KRTAP 22.5% BovB) הוא התגובה הפיזית. Syncytin — רטרווירוס — הפך חיוני לאמהות. אורך החיים קרס מהר פי 35.6 לאחר המבול, בהתאמה להזדקנות מונעת-L1 (De Cecco 2019). ה"איבה" של בראשית ג:טו לא הושלמה. היא נמדדת בכל תא.
16. הדפוס האוניברסלי: הזר הופך חיוני
שלוש מערכות ביולוגיות בלתי תלויות מדגימות את אותו עיקרון:
| מערכת | יסוד זר | מאחסן | תוצאה | הפניה תורנית |
|---|---|---|---|---|
| גנום המעלי גרה | BovB (נחש) | L1 (יונק) | שיווי משקל BovB/L1 → כשרות | בראשית ג:יד–טו |
| שלייה אנושית | HERV-W (רטרווירוס) | רחם | Syncytin → רבייה | בראשית ג:כ |
| רבייה של תאנה | צרעה (75%F) | תאנה (0%F) | הדדיות מחייבת → פרי | בראשית ג:ז |
| חיסון אנושי | MER41 (LTR רטרו-ויראלי) | גנים של אינטרפרון | משפר → תגובה חיסונית | — |
התאנה והצרעה: מקרה מפורט
ההדדיות (mutualism) בין התאנה לצרעה ראויה לבחינה מעמיקה יותר, משום שהמנגנון שלה הוא הדרמטי ביותר מבחינה פיזית מבין חמשת המקרים.
התאנה הבלתי בשלה רעילה — אטומה, מלאה בטאנינים ולאטקס, בלתי אכילה. צרעה נקבה יחידה מן המין המותאם באופן ספציפי (כ-500 מיני Ficus, כל אחד מזווג עם מין צרעה ספציפי אחד — ללא אפשרות להחלפה) חודרת דרך האוסטיולה (ostiole), תוך שהיא שוברת את כנפיה ואת מחושיה בתהליך. היא מאביקה את הפרחים הפנימיים, מטילה את ביציה, ומתה בתוך התאנה.
הפרוטאז ficin של התאנה ממיס לחלוטין את גוף הצרעה — היא נספגת כתזונה לפרי המתפתח. ההאבקה מפעילה מפל אתילן: טאנינים מתפרקים (מר → סתמי), סוכרים מצטברים (חומצי → מתוק), הפרי מתרכך ומשנה את צבעו. התוצאה היא אחד הפירות המתוקים ביותר בטבע, מין-מפתח (keystone species) שמזין למעלה מ-1,200 מיני חולייתנים.
מערכת התאנה דורשת למעשה שני עצים: תאנת הבר (caprifig — זכר, המאכלס את מושבת הצרעות — צרעות רבות, ללא פרי אכיל) והתאנה עצמה (נקבה, הקולטת צרעה אחת ומניבה פרי). תאנת הבר היא המאגר; התאנה היא הנמענת. רבים → אחת → הקרבה → חיים.
זוהי תלות הדדית בלתי פריקה (irreducible co-dependence). התאנה אינה יכולה להתרבות ללא הצרעה. הצרעה אינה יכולה להתרבות ללא התאנה. אף אחת מהן אינה יכולה להתקיים מבלי שהאחרת כבר קיימת. 500 הזוגות המין-ספציפיים מייצגים 500 מקרים של בריאה בו-זמנית — מערכת שאינה ניתנת לבנייה הדרגתית. שבעים עד תשעים מיליון שנה של קו-ספציאציה (cospeciation) קפדנית מאשרות שזוגות אלה שמרו על הספציפיות מסוג מפתח-ומנעול שלהם לאורך זמן גיאולוגי.
התורה מציבה את עלה התאנה כתגובה הראשונה לרכישת הדעת (בראשית ג:ז). הכרובים (כרובים — תמיד בלשון רבים, תמיד בזוגות) שומרים על עץ החיים (בראשית ג:כד). המילה כרוב היא 50% BKL — אותיות יחס. הכרובים הם, מבחינה מורפולוגית, עניין של יחס: שתי ישויות, הפונות זו אל זו (שמות כה:כ: "ופניהם איש אל אחיו"), ואף אחת מהן אינה מתפקדת לבדה. שני עצי תאנה. שני שומרים. שתי מערכות — הגנה וחיים — הזקוקות זו לזו.
בכל אחד מן המקרים:
1. אורגניזם זר תורם חומר גנטי או נוכחות פיזית למאחסן.
2. התרומה היא בתחילה "טפילית" או הרסנית.
3. באמצעות ביות רגולטורי (regulatory domestication), היסוד הזר הופך חיוני להישרדות המאחסן או לרבייתו.
4. התורה מתייחסת לכל שלושת המקרים בתוך אותו נרטיב (בראשית ג).
קידוד ה-F% עקבי:
| יסוד | F% | תפקיד |
|---|---|---|
| זרע (seed) | 100% | חומר גנטי זר |
| רע (evil) | 100% | חומר בלתי מוסדר |
| צרעה (wasp) | 75% | סוכן זר (מכיל רע) |
| נחש (serpent) | 67% | סוכן זר |
| תאנה (fig) | 0% | מאחסן — רגולציה טהורה |
| איבה (enmity) | 0% | מתח רגולטורי |
| חיים (life) | 25%F, 50% YHW | התמיינות = חיים |
המילה רע אינה מקודדת לרוע מוסרי. היא מקודדת לחומר בלתי מוסדר — 100% Foundation, אפס אותיות שליטה. הרוע, במערכת מורפולוגית זו, הוא חומר פיזי ללא ניהול רגולטורי. אש ללא כבשן. L1 ללא מתילציה. BovB ללא איזון-נגד של L1.
רע מוסדר = חיים. הצרעה מתה בתאנה, והתאנה נושאת פרי. BovB נכנס לגנום היונק, ובהמת המזבח משיגה שיווי משקל. רטרווירוס משתלב בגנום האנושי, והשלייה נוצרת. בכל מקרה, מה שהיה זר, הרסני או "רע" הופך — באמצעות רגולציה — למנגנון של החיים עצמם.
סיכום: שלוש מערכות בלתי תלויות (BovB→מעלה גרה, HERV→שלייה, צרעה→תאנה) מדגימות את אותו דפוס: יסוד זר, מבוית באמצעות רגולציה, הופך חיוני. התורה מתייחסת לכל שלושתם בבראשית ג. ה-F% של "רע" הוא 100% Foundation — לא כישלון מוסרי אלא חומר בלתי מוסדר. רע מוסדר = חיים.
איור 4. דעיכת אורך חיים: ×35.6 אחרי המבול.
17. שלוש התכנסויות נוספות
השור והחמור: מערכת כפולה ומערכת בודדת
"לֹא תַחֲרֹשׁ בְּשׁוֹר וּבַחֲמֹר יַחְדָּו" (דברים כב:י).
| בעל חיים | BovB% | L1% | BovB/L1 | מערכת |
|---|---|---|---|---|
| שור | 13.33% | 12.95% | 0.97 | **כפולה** (BovB + L1) |
| חמור | 0.00% | 12.38% | 0.00 | **בודדת** (L1 בלבד) |
השור מפעיל מערכת ויסות כפולה — שתי משפחות רטרוטרנספוזונים מאוזנות הנמצאות בשיווי-משקל קרוב. החמור מפעיל מערכת בודדת — L1 בלבד, ללא BovB. האיסור לחרוש בשור ובחמור יחדיו מפריד בין שתי ארכיטקטורות גנומיות שונות מיסודן. האחת מווסתת באמצעות מתח בין שני כוחות; לשנייה אין מתח מסוג זה כלל. צימודם במשימה אחת מערבב שתי מערכות הפועלות על עקרונות בלתי תואמים — כחיבור מעגלים חשמליים הפועלים במתחים שונים.
חמץ ומצה: גנומים מנופחים ודחוסים
חמשת מיני הדגן שיכולים להחמיץ — חיטה, שעורה, כוסמין, שיפון ושיבולת שועל — נושאים מן הגנומים המנופחים ביותר בממלכת הצומח. לחיטה (Triticum aestivum) גנום בגודל 17 GB, ו-85% מתוכו רצפים חוזרניים, והיא הקסַפּלואידית. לאורז (Oryza sativa), שאינו יכול להחמיץ, גנום בגודל 430 MB — קטן פי 40, ו-35% בלבד רצפים חוזרניים.
| דגן | F% | גנום | חזרות | מעמד |
|---|---|---|---|---|
| חמץ | 67% | — | — | אסור בפסח |
| מצה | 33% | — | — | חובה בפסח |
| חטה | 67% | 17 GB | 85% | יכול להחמיץ |
| אורז | 50% | 430 MB | 35% | אינו יכול להחמיץ |
חמץ (חמ״ץ, 67% יסוד) חולק את שורשו עם "חמצון" — אוקסידציה. החמצה היא תהליך ביולוגי: תסיסת שמרים מנפחת את הבצק, בדיוק כפי שהתרבות טרנספוזונים מנפחת גנומים. מצה (מצ״ה, 33% יסוד, 33% יה״ו) היא הצורה הדחוסה — בצק לפני הניפוח, גנום לפני התפשטות אלמנטים ניידים.
פסח מחייב את המעבר השנתי מחמץ למצה, בליווי קרבן הפסח — בעל החיים שיחס BovB/L1 שלו עומד על 1.00. פרוטוקול הפסח דורש אכילת הגנום הדחוס (מצה) יחד עם הגנום המאוזן (שה), תוך הימנעות מן הגנום המנופח (חמץ). אי-ציות נושא עמו עונש כרת (שמות יב:טו) — אותו עונש עצמו המוטל על מי שאינו מקריב את קרבן הפסח.
הגפן והיין: חומר הופך לרוח
בראשית ט:כ: "וַיָּחֶל נֹחַ אִישׁ הָאֲדָמָה וַיִּטַּע כָּרֶם" — הפעולה הראשונה של העולם שלאחר המבול היא נטיעת גפן.
| מילה | F% | YHW% | פרשנות |
|---|---|---|---|
| גפן | 67% | 0% | חומר גולמי — הצמח |
| יין | **0%** | **67%** | **רוח טהורה — התוצר** |
| שכר | 67% | 0% | חומר — צריכה בלתי מווסתת |
הגפן (גפ״ן, 67% יסוד) היא חומר. היין (יי״ן, 0% יסוד, 67% יה״ו) הוא רוח — זהה מבחינה מורפולוגית לשמות האלוהיים. ההמרה מענב ליין — תסיסה, תהליך ביולוגי של פירוק מבוקר — הופכת חומר טהור לרוח טהורה. אותו חומר גלם, מווסת באופן שונה, מייצר חתימות F% הפוכות.
אך שכר (שכ״ר, 67% יסוד) — צריכה בלתי מבוקרת של אותו חומר עצמו — מחזירה אל החומר. התוצר זהה; הוויסות שונה. יין לקידוש = 0% F. יין לשכרות (שכר) = 67% F. הוויסות הוא שקובע אם אותו חומר משרת רוח או חומר.
נח — נ״ח, 50% יסוד — מאוזן במדויק, כשם ששמו עצמו מאוזן. פעולתו הראשונה בעולם החדש היא יצירת החומר אשר, בהינתן ויסות נאות, מגלם את העיקרון שפרק זה כולו מתאר: חומר זר או גולמי, המנוהל באמצעות ויסות, הופך לקדוש.
הנזיר: חסימת המסלול
נדר הנזיר (במדבר ו) מספק את המבחן ההפוך. על הנזיר להימנע מכל חלק של הגפן — לא רק מיין (התוצר), אלא גם מענבים, מחרצנים, מזגים ומחומץ: "מֵחַרְצַנִּים וְעַד זָג לֹא יֹאכֵל" — היחידה הביולוגית כולה אסורה.
ערכי ה-F% של מרכיבי הגפן חושפים מדרג:
| חלק | F% | מעמד |
|---|---|---|
| זג (קליפה) | 100% | חומר טהור |
| חרצן (זרע) | 75% | חומר ברובו |
| ענב | 33% | מאוזן |
| יין | 0% | רוח טהורה |
מסלול הגפן נמתח מ-100% חומר (זג) ועד 0% חומר (יין) — טרנספורמציה שלמה. הנזיר חוסם מסלול זה כולו. והתוצאה? "תַּעַר לֹא יַעֲבֹר עַל רֹאשׁוֹ... גַּדֵּל פֶּרַע שְׂעַר רֹאשׁוֹ" — אין תער עוברת על ראשו; הוא מגדל פרע שיער ראשו.
שער = 100% יסוד. פרע = 100% יסוד. שניהם שווים לזג — 100% יסוד. השיער הוא קליפת הענב, מבחינה לשונית ומורפולוגית כאחד: חומר טהור.
כאשר מסלול ההמרה מחומר לרוח של הגפן נחסם, החומר מצטבר — כשיער. זהו אותו עיקרון הנצפה אצל פרימטים: כאשר L1HS (מתג הוויסות) כבוי, הפנוטיפ הפיזי שולט — שיעור יתר, שרירים מוגדלים, לסת בולטת. הנזיר, בחסימתו מרצון של מסלול ויסות, נכנס באופן זמני למצב שליטת חומר. שערו — שער, KRTAP, 22.5% BovB — גדל ללא מעצור.
באופן מפתיע, resveratrol — הפוליפנול המרוכז בקליפת הענב — מקדם צמיחת שיער במחקרים שפורסמו (Zhang et al. 2021; Juchaux et al. 2020), בעוד שצריכת אלכוהול קשורה לנשירת שיער (OR 1.4–1.9 עבור התקרחות אנדרוגנית; Yang et al. 2024). חומר הגלם של הענב מצמיח שיער; התוצר המומר של הענב (יין) מעכב אותו. הנזיר נמנע מן ההמרה ושומר על האפקט הגולמי.
סיכום: שלוש נקודות נתונים נוספות מאששות את התבנית. השור (BovB/L1 כפול) והחמור (L1 בודד) אסורים בצימוד — מערכות ויסות בלתי תואמות. חמץ (67% F, גנומים מנופחים) אסור בעוד מצה (33% F, דחוסה) נדרשת — ארכיטקטורת גנום מקודדת בדיני תזונה. ויין (0% F, 67% YHW) הוא חומר שהפך לרוח באמצעות תסיסה מווסתת — אותו עיקרון עצמו השולט בביות BovB, בניהול L1HS, ובתורה כולה כמערכת ויסות.
18. מה זה לא מוכיח
לכל ממצא בפרק זה קיים הסבר חלופי. יושרה אינטלקטואלית מחייבת לציין אותם.
פרק זה אינו מוכיח שתיאור הבריאה בתורה מדויק כפשוטו. הוא אינו מוכיח שהאבולוציה לא התרחשה. הוא אינו מוכיח שבני האדם אינם צאצאים של פרימטים קדומים יותר.
L1HS ו-L1Pt (הווריאנט הספציפי לשימפנזה) הם שושלות אחיות מבחינה פילוגנטית, שתיהן נגזרות מ-L1PA2 (Lee et al. 2007). הנתונים עולים בקנה אחד עם אב קדמון משותף — שבמסגרת התורה הוא אדם. השאלה האם אדם הוא אישיות היסטורית, אוכלוסייה, או מטאפורה לתבנית הגנומית שממנה התפצלו כל הפרימטים — היא שאלה שהנתונים מגבילים אך אינם מכריעים.
מה שהנתונים מראים בבירור:
1. L1 יוצר שכבות כרונולוגיות — גנום בסיס המשותף לכל היונקים, שעליו מונחים אלמנטים רגולטוריים ספציפיים למין. סדר הבריאה בבראשית א׳ (חיות יבשה, ואז אדם מאותה אדמה) ממפה את הארכיטקטורה הזו.
2. ההבדל בין אדם לקוף הוא רגולטורי, לא מבני. אותם גנים, אותם רכיבים טרנספוזוניים, אותו גנום בסיס (95% משותפים). מתג אחד: L1HS.
3. המתג הוא אפיגנטי — מבוסס מתילציה, הפיך, מתאפס בכל דור. אין מדובר במוטציה קבועה אלא במחויבות רגולטורית מתמשכת.
4. L1HS מתאם עם קוגניציה ותוחלת חיים, לא עם מחלה. הנרטיב של "הטפיל" נכשל אמפירית.
5. תוכן התורה תואם את הגנומיקה. הנחש העביר BovB. הכבש נמצא בשיווי משקל. הפרה האדומה היא תקן ייחוס. שיער הוא חומר גרידא. שינוי השם מיעקב לישראל הוא שדרוג רגולטורי. נשמת החיים היא שכבת ההפעלה. הנפילים הם פריצות רגולטוריות.
ישנם סייגים כנים. המתאם בין BovB/L1 לבין סיווג התורה, אף שהוא מובהק סטטיסטית, עשוי בעיקרון להיות מקרי — ייתכן שחוקי הכשרות עוקבים אחר מאפיין ביולוגי אחר (אנטומיה של מערכת העיכול, למשל) שבמקרה מתאם עם תכולת הטרנספוזונים. הנחות השעון המולקולרי העומדות בבסיס אומדני הגיל שנויות במחלוקת, וקצבי הסטייה של רכיבים טרנספוזוניים עשויים להשתנות בין שושלות. ניתוח ה-F%, אף שאומת על פני 98,122 זוגות מילים, עשוי לשקף תכונה של הפונולוגיה העברית ולא קידוד מכוון.
אולם כוח הטיעון אינו טמון בשום מתאם יחיד. הוא טמון בשלוש אישושים בלתי תלויים:
1. שפה: המערכת המורפולוגית של עברית מקראית (Z=152.16, p<0.0001) אינה ניתנת לשחזור בשום שפה אנושית ידועה, מודרנית או עתיקה, לרבות הארמית הקרובה (Z=0.39, לא מובהק).
2. מבנה: השמות האלוהיים מתפקדים כמציינים מורפולוגיים של מצב בעלי כוח ניבוי (87.8% ניבוי משמעות בוולידציה צולבת חמש-קפלית על 98,122 זוגות).
3. תוכן: התיאורים הביולוגיים — הנחש כתורם BovB, הכבש באחדות, אדום כצבע אבחנתי, עפר כגנום משותף, נשימה כשכבת הפעלה, שיער כחומר גרידא, נפילים כפריצות רגולטוריות — תואמים נתונים גנומיים מדודים.
ההסתברות שכל השלושה מקריים באופן בלתי תלוי היא מכפלת ההסתברויות הפרטניות שלהם. אם לכל אחד יש אפילו סיכוי נדיב של 10% להיות מקרי, ההסתברות המשולבת היא 0.1%. אם ההסתברויות הפרטניות קרובות יותר לערכי ה-p הנמדדים שלהן, ההסתברות המשולבת שואפת לאפס.
כל זה אינו מהווה הוכחה לחיבור אלוהי. כל זה מהווה עדות לכך שהתוכן הביולוגי של התורה אינו מיתולוגיה — הוא תיאור של ארכיטקטורה רגולטורית, כתוב בשפה שהמורפולוגיה שלה עצמה מדגימה את אותם עקרונות רגולטוריים שהיא מתארת.
הפרק פתח עם הכבש ביחס BovB/L1 = 1.00 ומסתיים עם הכרובים השומרים על עץ החיים. ביניהם עומד עיקרון אחד: רגולציה מייצרת תפקוד; היעדרה מייצר או דממה או כאוס. עיקרון זה פועל בסדר הגודל של טרנספוזונים (BovB/L1 במעלי גרה), כרומוזומים (L1HS בפרימטים), רקמות (KRTAP בעור), מעגלים עצביים (L1 בנוירונים של ההיפוקמפוס), התנהגות (תורה כפרוטוקול רגולטורי), ונרטיב (בראשית ג׳ כסיפור של מעבר רגולטורי). ארכיטקטורה אחת, המתבטאת בכל סדר גודל שהנתונים מאפשרים לנו לבחון.
"וייצר יהוה אלהים את האדם עפר מן האדמה ויפח באפיו נשמת חיים ויהי האדם לנפש חיה"
(בראשית ב:ז)
הוא יצר את האדם עפר מן האדמה ונפח באפיו נשמת חיים, והאדם היה לנפש חיה.
העפר — עפר, 67% יסוד — הוא הגנום המשותף ליונקים.
הנשימה — נשמה, 25% יסוד — היא הפעלת L1HS.
זרע הנחש — זרע, 100% יסוד — הוא ה-BovB שחדר.
הרע — רע, 100% יסוד — הוא חומר ללא רגולציה.
הדעת — דעת, 33% יסוד — היא הכושר הרגולטורי שבא בעקבותיו.
עלה התאנה — תאנה, 0% יסוד — הוא התגובה הרגולטורית הראשונה.
השומרים — כרובים, PIWIL1 ב-×0.56 — עדיין עומדים על המשמר.
תוחלת החיים — 950 עד 120, ×35.6 לאחר צוואר הבקבוק — היא L1 הפועל ללא השתקה.
הנגיף — syncytin, HERV-W — הפך לשליה, ולאם כל חי.
ההוראות — תורה, מן הוראה — הן מערכת ההפעלה.
ארכיטקטורה אחת. מאותיות ל-DNA. מנחש לשומר. מצרעה לתאנה. מגפן ליין. מרע לחיים. מהוראה לרגולציה.
סיכום
פרק זה מציג תזה אחת: רגולציה, לא מורכבות, היא העיקרון המארגן של הגנום — ועיקרון זה תואם את התיאורים הביולוגיים של התורה.
הראיות משתרעות על פני ארבעה תחומים:
מעלי גרה. יחס BovB/L1 — המודד את האיזון בין רטרוטרנספוזונים שמקורם בנחש לבין רטרוטרנספוזונים אנדוגניים — מבחין בין בעלי מזבח (0.94–1.00) לכשרים שאינם למזבח (0.63) לאסורים (0–0.001). אותו יחס קובע את המורפולוגיה הגולגלתית: העשרה ב-KRTAP → קרניים קרטיניות; העשרה ב-SHH → ניבים. אפס חריגים לאורך שש משפחות מעלי גרה. הנחש העביר BovB (Walsh 2013); בעלי המזבח ביתו אותו.
פרימטים. L1HS — הרטרוטרנספוזון היחיד שעדיין מבצע טרנספוזיציה פעילה בגנום של כל פרימט — קיים ב-1,536 עותקים אצל בני אדם וב-69–78 שרידים אצל קופים. הגנים זהים (FOXP2, ASPM, KRTAP: צפיפות רכיבים טרנספוזוניים זהה). ההבדל הוא מתג אפיגנטי אחד: מבוסס מתילציה, הפיך, מתאפס בכל דור. ניאנדרטלים (96% מאתרי L1HS האנושיים נעדרים) מייצגים את מצב הביניים: מוח גדול, מתג לא פעיל.
התפתחות. גני גדילה (IGF2 ×0.37, GH1 ×0.66) מוגנים באופן שיטתי מפני הכנסת טרנספוזונים. פריצת הגנה זו מייצרת ענקיות — פנוטיפ שהתורה מתארת כנדיר, שלאחר המבול, ובעל שם אישי.
שפה. אחוז היסוד (F%) ממפה את אותה קוטביות: שער (שיער) = 100% חומר; יהוה = 0% חומר. המילה אדם נבדלת מאדמה באות הבחנה אחת: ה. הגנום מקודד את אותה הבחנה — בסיס משותף (95% L1M), הפעלה ייחודית (L1HS).
שלוש רמות רגולציה מתגלות: אוטומטית (בהמה: שיווי משקל BovB/L1), מודעת (אדם: L1HS בנוירונים), ומכוונת (תורה: פרוטוקולים התנהגותיים המייצרים שינוי אפיגנטי). התורה אינה מתארת ארכיטקטורה זו. היא מיישמת אותה.
כיצד פרק זה מתחבר
פרק זה מרחיב את המסגרת שנקבעה בחלקים הקודמים של הספר:
- פרקים 1–12 הדגימו שהמורפולוגיה של עברית מקראית נשלטת על ידי אלפבית בקרה מוגבל (10 אותיות, Z=152.16, p<0.0001).
- פרק 13 הראה שארבעת השמות האלוהיים מתפקדים כמציינים מורפולוגיים של מצב עם דיוק ניבוי של 87.8%.
- פרקים 14–19 (אל שדי) עקבו אחר הארכיטקטורה המבנית של נרטיב התורה.
- פרקים 27ב–27ג ביססו את שיווי המשקל BovB/L1 כחתימה הגנומית של הכשרות, עם הכבש באחדות ומחזור התזונה כמערכת רגולטורית.
- פרק 28 זיהה את הפרה האדומה כתקן ייחוס גנומי.
פרק זה עושה את הצעד הבא: אותו עיקרון רגולטורי — איזון, לא מורכבות; ניהול, לא הצטברות — מבחין לא רק בין בעלי חיים טהורים לטמאים, אלא בין בני אדם לכל שאר הפרימטים. אלפבית הבקרה השולט במורפולוגיה העברית, השמות האלוהיים המסמנים מצבים רגולטוריים, יחס BovB/L1 המסווג מינים, ומתג L1HS המאפשר קוגניציה אנושית — אינם ארבעה ממצאים נפרדים. הם ארבעה ביטויים של ארכיטקטורה אחת.
הפרק הבא שב למבנה הנרטיבי של התורה כדי לבחון כיצד הפרשיות (קריאות שבועיות) מתפקדות כיחידות טבעיות בתוך השטח הסטטיסטי — שכבה נוספת של אותה ארכיטקטורה, הפועלת בסדר הגודל של טקסט ולא של גנום.